Theoretical Biology
Translated from: Э. С. Бауэр, Теоретическая биология, Издательство ВИЭМ, Москва–Ленинград, 1935.
Translation note: A complete English translation of Bauer’s 1935 treatise, made directly from the Russian. Foreign terms are glossed on first use as [Cyrillic — transliteration]; the author’s own notes appear as sidenotes marked “[Author’s note:]”. Mathematical formulae, tables, and figures are reconstructed from the original; passages where the 1935 source is inconsistent or misprinted are reproduced faithfully and flagged in translator’s sidenotes.
Preface
The book here offered has a history of development of more than sixteen years. The first stage of this history was the little book “Grundprinzipien der rein naturwissenschaftlichen Biologie”, which appeared in 1920; the second stage was the book “Physical Foundations in Biology”, which appeared in 1930. Now, five years on, the present book is to constitute the third stage.Bauer’s two earlier works: Grundprinzipien der rein naturwissenschaftlichen Biologie (“Fundamental Principles of a Purely Scientific Biology”), 1920, written in German; and Физические основы в биологии (Fizicheskie osnovy v biologii, “Physical Foundations in Biology”), 1930.
Although between the first two books there lies an interval of ten years, and between the last two only five, the difference in content is in the second case considerably greater than in the first.
And yet the development can by no means be regarded as complete. The experimental material is rapidly increasing, and could no longer be presented in full even in this book.
As regards the exposition, the book constitutes a logical whole, and not a single proposition can be correctly understood out of connection with the rest. For this reason one cannot obtain a correct conception of the treatment of any given question by reading only a single particular chapter. On the other hand, the demonstrations of the various propositions touching, for example, stimulation, the regularities of evolution, metabolism, the structure of living matter, and so forth, are by no means exhausted by the experiments and facts adduced in the corresponding chapters. These propositions are equally supported by facts adduced in another connection in other places.
As regards their substantiation by facts, it must be borne in mind that the general and fundamental principles which we here expound are founded not only upon the facts adduced in the book, but also upon a great quantity of known facts, which, after all, cannot all be adduced.
That the preservation of non-equilibrium in the thermodynamic respect is a characterising property of living systems — this is the conclusion, for example, to which all investigators have come who have occupied themselves more closely with the significance of the resting metabolism. The factual material that compelled these investigators to arrive at such a conclusion could not, naturally, be set out here.
Thus, for example, A. V. Hill writes: “The living cell represents a complex organised system …, which in the thermodynamic respect is infinitely improbable, and can remain in this state only so long as free energy can be employed for the maintenance of this organisation.”[Author’s note:] A. V. Hill, Supplement to Nature, No. 3106, May 1929.
Thus, too, I. Straub comes to the conclusion that “the analogy in the relationship of yolk and white, of blood cells and serum, of yeast cells and nutrient solution, of algae and the surrounding medium, makes it necessary to consider systematically the deviation in respect of equilibrium as a primary property of independent living beings.” Such pronouncements may be found in many other investigators. The more surprising it is that, while acknowledging this proposition, they do not draw from it all the conclusions, and firmly adhere to the conception of a constant disturbance of equilibrium from without. The task, therefore, consisted not only in the substantiation of the principles by facts, but also in the theoretical substantiation of the facts themselves.[Author’s note:] I. Straub. Cited after Berichte über die gesamte Physiologie. Abt. B, Bd. 77, H. 7, May 1934.
There is no doubt that in the majority of the chapters there are deficiencies and errors. For every factual correction, and for the indication of every fact that might contradict the corresponding theoretical propositions or confirm them, I should be exceedingly grateful to the specialists working in the corresponding fields.
In the reading of proofs and the editing of the text I was helped by many collaborators, in particular by my collaborator V. S. Brandgendler, to whom I here express my gratitude. I am especially indebted to my wife S. S. Bauer, who has gone the whole way of this work from its very beginning, helping me especially in the physical and mathematical side of the matter, checking the calculations, preparing the drawings, and so forth.
In the main, however, I owe the writing of this book to the unfolding of the construction of our socialist motherland, which has set before science the task of a deepened scientific substantiation of the practical measures in agriculture and medicine, and which secures the ideological and material support for this difficult work.
E. Bauer, 27 April 1935.
Part I: General Theory of Living Matter
Introduction: Subject and Method of Theoretical Biology
Biology is the science of life, or more correctly of living beings. It is the science of the laws of motion [движение — dvizhenie] (in the very broadest sense of the word) of organised living matter [организованная живая материя — organizovannaia zhivaia materiia].движение (motion) is used throughout in the dialectical-materialist sense: it encompasses all change and development, not merely physical displacement. “The laws of motion of living matter” is Bauer’s definition of biology.
In view of the boundless diversity of forms and functions of living organised matter which we find in living beings, it becomes understandable that, first of all, it was necessary to describe and bring into order this diversity of forms and functions, and also to establish certain regularities [закономерность — zakonomernost′] of it.закономерность (regularity) ≈ German Gesetzmäßigkeit, “law-governed regularity.” It denotes the observable, phenomenal conformity-to-law of a process, and is kept strictly distinct throughout from закон “law” [закон — zakon], the underlying general law of which a regularity is the manifestation.
Here attention was at first directed chiefly to those manifestations of life which can be observed and described under natural conditions (such as reproduction, growth, metamorphosis, mode of life). On the basis of similarity in the structure and functions of certain organs, for example the sexual organs, and in the manner of reproduction, living beings were combined into groups, and thus, with the further deepening of knowledge concerning the structure of organisms, there arose the concept and delimitation of species. But the scientific, theoretical foundation for this combination and delimitation of various organisms according to their structure and mode of life was obtained only in the theory of evolution, established and elaborated by Darwin, according to which all the manifold forms, the various species, arose one from another. Only the recognition that all the various existing species possess various degrees of “blood kinship” with one another made possible the construction of a genealogical tree of species, on the basis of which it became possible both to explain regularities already found and to discover new ones. The theory of evolution had just as decisive a significance for the deepening of knowledge concerning the regularities of the various forms of the above-mentioned vital phenomena, such as reproduction, metamorphosis and others, and also for the doctrine—developed during this same time—of the arising and preservation of new forms, that is, for that doctrine, closely bound up with evolution, of the inheritance of properties, which obtained its scientific foundation in the laws of heredity discovered by Mendel. Thus there developed, as the principal branches of biology, morphology as the science of the forms and structure of living beings, embryology as the science of the individual development and formation of forms and organs in the individual, the doctrine of heredity and of the origin of species, or the doctrine of the history of the origin of living beings. As concerns such vital manifestations of functions as reproduction, fertilisation, mode of life (manner of nutrition, general conditions of existence, and so forth), even at the present time there is understood by biology the study of these vital functions, susceptible of observation and description under natural conditions. The various, exceedingly manifold forms of these vital manifestations obtained their theoretical natural-historical explanation on the basis of evolutionary theory, in connection with the explanation of the diversity of structure and with the descriptive history of the development of the various living beings.
Thus, evolutionary theory is that theory to which the material accumulated by descriptive biology concerning the diversity of forms and the vital manifestations of organisms observed under natural conditions has led, and which became the guiding principle of these investigations.
Gradually, however, it became ever clearer that the description of structure, development and vital manifestations ultimately reveals only the regularities of the manifestation of living matter, only the result of its laws of motion, and not these laws themselves; from this it follows that evolutionary theory is a theory of the manifestation of living matter in the course of history, that it takes into account only a part of the vital phenomena and does not represent a theory of the law of motion of living matter which gives rise to these phenomena. Evolutionary theory is thus the theoretical culmination, the greatest scientific achievement of the first, descriptive period of biology. When we designate evolutionary theory as the theoretical culmination of the first, descriptive period of biology, this is in no way contradicted by the fact that for the foundation of this theory there also served a rich experimental material (especially that of animal- and plant-breeders). For the content and significance of a theory, the decisive thing is not the methods of its foundation, but the phenomena to be explained.
Corresponding to this, there developed ever further those directions and branches of biology which studied the laws of motion of living organised matter that find their expression in the various observable vital manifestations and forms. This led to the development of physiology, the mechanics of development, genetics—that is, of the experimental sciences—owing to the striving to analyse those particular acting forces or motions and changes within the organism which give rise to the given vital manifestations. Thus, physiology studies the functions and mechanism of the particular organs making up the organism as a whole, their exchange of matter and energy, the laws by which they react to external influences, that is, their irritability; the mechanics of development investigates those factors, or conditions, which determine the formation of forms and functions during embryonic development, whereby, also making use of experimental methods, it acts in a planned manner upon the course of this development and alters or excludes determinate conditions. Thus, modern genetics strives, by means of systematic crossings and the simultaneous cytological investigation of chromosomes, to derive the observable laws of heredity from those laws by which the reactions and changes of the hereditary mass located in the sexual cells take place. Parallel with the development of these sciences and with the deeper penetration into the particular processes and their mechanism—that is, with the elucidation of the regularities and forces operating therein—ever greater attention was directed to the physical and chemical phenomena participating in these processes, to the study of these phenomena, whereby attempts were even made to reduce particular vital processes in the organism exclusively to them.
Here we must point to the following. The essential distinction of sciences such as physiology, the mechanics of development, genetics, and in general those which we unite under the name of experimental biology, lies by no means in their experimental character (as is often depicted), in contrast to the earlier branches of biology, such as comparative morphology and descriptive biology. The essential distinction of these newly developed divisions of biology lies in the new and deepened posing of the question and in those new tasks which these sciences set themselves. Herein the experimental method is the natural, but by no means the sole and decisive means for the resolution of these tasks. The essential distinction and the novelty of the tasks set by these sciences themselves consisted precisely in the striving to find the laws of motion of living matter and with their aid to explain the regularities of the various forms of their manifestation under various conditions. These sciences set themselves the task of studying those laws of motion of living matter which are proper to it itself and which manifest themselves of necessity in all vital phenomena. Just as the resolution of the first task—the finding of regularities in the forms of the manifestation of living matter—inevitably had to begin with the description of particular forms and vital phenomena, their classification and systematisation, so too, just as naturally, the resolution of the second task had to begin with the description and finding of the individual laws of motion of living matter underlying the various individual manifestations of life and processes of form-formation.
Thus there developed the various special divisions of physiology, such as the doctrine of the exchange of matter and energy, which broke up into separate branches corresponding to the various energy-supplying processes, such as the doctrine of respiration, of fermentation, of nitrogenous, fatty, and intermediary metabolism, of syntheses in the organism; further, the doctrine of the reactions of the organism and of its irritability, which broke up into the doctrines of muscular contraction, of the conduction of stimulation together with the chemical and physical phenomena accompanying these processes; the doctrine of reflexes, and so forth. The like we observe also in embryology, where the mechanics of development founded by W. Roux (or, as it is now often called, the physiology of development) subjected to analysis the particular, especially the very earliest stages of development, investigating the various fates of particular cells, and the factors determining this fate, including the chemical and physical phenomena. The very same we see also in genetics, which in its modern form has been transformed chiefly into an experimental investigation of the structure of chromosomes, its changes, and so forth, in order in this way to derive the external manifestation of characters and the regularities of their inheritance from the laws of motion and acting forces proper to the bearers of these hereditary characters.
Thus, Darwin’s theory gave a general principle for explaining the diversity of forms and functions of organisms as they appeared on the earth’s crust—a theory embracing the regularities of the forms of living beings, of their origin and of their law of motion for all living organisms; now the question arises whether such a theory is also possible for every living organised matter independently of the historical conditions of its development. In other words, is it possible to find such universal laws of motion of living matter as are valid in all its forms of manifestation, however manifold these forms might be—that is, do there lie at the basis of all the various laws of motion which have already been found, or are yet to be discovered, by the particular biological disciplines in the field of genetics, physiology, the mechanics of development, and so forth—do there lie at their basis such universal laws of motion of living matter, the various manifestations of which constitute these particular laws of the named special divisions of biology, just as the special, particular laws of the descriptive sciences—morphology and descriptive biology—constitute the various manifestations of the universal regularities of evolution. But this question is equivalent to the question whether living matter has its own peculiar laws of motion. And since we have defined biology as the science of the laws of motion of organised living matter, this question is analogous to the question whether there exists a science of life, biology, or whether it is merely a division of applied physics and chemistry. For if we ascribe to living, organised matter its own laws of motion, proper precisely to living matter and to it alone, constituting its attribute, the form of its existence, then these laws must come to light in every form of the manifestation of living matter. Then all the special laws of motion which have been and will be discovered by the particular branches of biology, such as the regularities of physiology, of the mechanics of development, of genetics, and so forth, must turn out to be special cases, special manifestations of the universal laws of motion proper to living matter, even though these manifestations should occur under historically conditioned and essentially altered conditions, and therefore also in an essentially altered form. There are only two possibilities of evading the inevitability of this conclusion.
-
We may assert that living matter has no laws of motion of its own, that the laws of motion of living matter are in essence the same as those of the non-living. Then, being consistent, we have no right to speak of living matter at all. But then biology is nothing other than applied physics and chemistry, that is, the application of the laws of physics and chemistry to those complex systems which living beings are.
-
Or else we assert that living beings have their own peculiar regularities, their own laws of motion, but that these laws are conditioned not by the state, organisation, structure of the matter of living beings—that is, are not a form of the manifestation and existence, an attribute of living matter—but are the result (and this conclusion is then inevitable) of supra-material, divine forces which do not alter the laws of physics and chemistry, but constantly direct them.
The first of these points of view is defended by the direction which is known in biology under the name of mechanism; the second by vitalism. Both directions halt before reaching the decisive point and endeavour to cover this halt with a false philosophy. But scientific investigation does not stand still and, despite the retarding influence of these two directions, penetrates ever deeper and deeper into the special and general regularities of the motion of living matter. There exists, of course, only one correct path to prove the inadmissibility of both indicated points of view, which deny the existence of general laws of motion proper to every living matter and to it alone, as the form of existence of this organised, living matter: this path consists in the finding and investigation of the general laws of living matter, in bringing to light how they manifest themselves in various forms in the various forms of living beings, and, on the other hand, in showing that in all the various particular regularities there manifest themselves one and the same general regularities of living matter, that in the manifold regularities of vital phenomena—changing in the course of the development of living beings on earth—there find their expression one and the same laws of motion, proper to living matter and to it alone.
Thus, we arrive at the conclusion that if we ascribe to living organised matter special laws of motion proper to it alone—that is, if we speak of a special science, biology—and at the same time wish to remain on the ground of materialism, then we must not only give an affirmative answer to our question whether it is possible to find such general laws as lie at the basis of all the laws of motion in the particular, special domains of biology, or of which these special regularities of physiology, the mechanics of development, genetics, and so forth, serve as particular concrete manifestations, but we must also say that to find these laws, to carry out these generalizations, and to apply their results as a guiding theory in investigation, is the pressing immediate task of theoretical biology.
On the basis of all that has been said above, we may form for ourselves the following schematic representation of the development of biology, of its special, experimental-descriptive, and its general-theoretical part:
| Special or descriptive biology | General or theoretical biology |
|---|---|
| Zoology, botany, morphology, descriptive embryology, descriptive biology, ecology, and so forth. | Evolutionary theory, or the doctrine of the origin of species. |
| Physiology, the mechanics of development, genetics, the investigation of protoplasm. | General theory of living matter. |
Of course, each point of this scheme stands in need of explanation.
In the first place, we equate the special with the descriptive-experimental, on the one hand, and the theoretical with the general, on the other. It seems to us that this equation is justified and in the main correct. For the theoretical treatment of any phenomenon consists precisely in this, that from the great diversity of special, concrete phenomena we abstract the general, the law-governed, and investigate this particular abstracted regularity, analyse it and bring to light its various possible forms. We assume determinate conditions and determine how the process must proceed under the given conditions, in what form the regularity will manifest itself under the given concrete conditions. Then we compare the result thus obtained with the data of experience, investigating how these phenomena proceed in nature there where the conditions we have assumed are realised; or else we ourselves create these conditions in experiment and investigate whether the process really proceeds as we have deduced it from the general law abstracted by us under the conditions we have assumed. If the general law thus abstracted is really confirmed—that is, if the conclusions drawn from it under the assumption of various conditions are corroborated in experience or in experiment, if our law is really a general regularity—then we can, with its aid and on the basis of an analysis of the concrete conditions given in experience, predict the course of the processes, or else, by means of a corresponding alteration of the conditions, direct them in the desired direction; that is, then this abstract general law becomes the property of theoretical science. It is in this sense that we speak of theoretical science in opposition to experimental or descriptive science. We speak of theoretical science when we proceed from the aggregate—obtained on the basis of all the experience of the given science—of purely abstract, general laws of the given domain of phenomena, and represent the phenomena observed in experience as particular cases of these laws under determinate conditions. Of descriptive or experimental science, on the contrary, we speak when, proceeding from observations and particular experimental results, we collect the factual material for theoretical science, or, generalising it, arrive at general regularities.
At first glance it may seem that the distinction here reduces only to the method of exposition. But this notion is entirely incorrect, since, in the first place, experimental, descriptive science differs from theoretical not only in how it represents facts and general regularities: besides this, the laws of experimental, descriptive science are derived in theoretical science in an entirely different way, so that the distinction between the two sciences lies not only in the method of exposition, but also in the method of investigation; and, in the second place, the laws obtained by theoretical science are, in their content too, not equivalent to the laws obtained by experimental-descriptive science by way of generalizations.
As concerns the first distinction, descriptive-experimental science obtains its laws by way of the generalisation of observations and experimental data, but the law thus derived is by no means yet a theory of the given group of phenomena and therefore by no means constitutes the content of theoretical science. This law becomes the content of theoretical science only when it can be deduced from determinate general laws and theoretical representations as a particular case for those conditions under which the law was found. Then and only then do we speak of a theory of the given phenomena. Thus, experimental science obtains its laws from experience by way of generalisation, whereas theoretical science obtains these same laws on the basis of general abstract propositions and assumptions, whereby determinate concretising conditions are adopted. Of course, these abstract laws and assumptions are also abstracted from experience and adopted on the basis of it, but they go beyond the bounds of the immediate data of experience.
Let us elucidate this difference with a few examples.
As the first let us take the law of the fall of bodies from mechanics. In textbooks of experimental physics this law is presented as the result of the generalisation of systematically conducted observations and experiments and measurements of the path traversed by falling bodies in various intervals of time. Galileo did indeed obtain this law precisely in this way, and since the time of Galileo the path of the systematic causal-analytical experiment has been considered the model of experimental investigation in biology too. It was of this model that W. Roux thought when founding the mechanics of development. To it Hartmann too points, in his “General Biology”, as the path that will lead biology forward:
“The great merit of Galileo consists in this, that he introduced the systematic experiment and thereby discovered a new kind of induction, which, having led with the inevitability of logical necessity to causal cognition, at the same time proved in the highest degree creatively fruitful for the advancement of science.”[Author’s note:] M. Hartmann, General Biology, I, 7. The law obtained in this way states, as is well known, that every freely falling body (if the resistance of the air may be neglected) falls with a constant acceleration and that the path traversed is equal to the half-product of this acceleration and the square of the time, that is
where is the path traversed, the acceleration, and the time.
But this very same law is obtained in theoretical mechanics by a different path: not on the basis of the generalisation of observations, but as a particular case of much more general propositions. These universal propositions are the Newtonian laws of motion and his law of the attraction of masses. As is well known, the latter states that the force with which two masses are attracted is proportional to these masses and inversely proportional to the square of the distance, that is
where is the force of attraction, and the two masses, their distance from one another, the factor of proportionality, or the force itself in the case of unit masses at unit distance.
Newton’s second law of motion states that force is equal to the product of mass and the acceleration of the moving body, that is
From these general laws there is derived in theoretical mechanics the Galilean law of fall as a particular case. Namely, if we denote the mass of the falling body by , the mass of the earth by , then we obtain
or
and since, in comparison with the radius of the earth, the change of distance may be neglected—that is, may be regarded as constant—we have:
that is, in falling the acceleration is constant, and the path traversed in the time we obtain from this formula by means of a double integration with respect to , which leads us directly to the law of fall
How far we may already speak here of a theory of the law of fall, and whence, by what path, these general laws of motion were obtained—of this we shall speak further on, when we investigate several examples from various domains. In any case, we see that the law of fall obtained by the second path also gives more in its content than the law obtained by way of the generalisation of analytical experiments and observations. Thus, for example, it shows that this is an approximate law, that it is valid only in so far as the length of the path traversed in falling may be neglected in comparison with the radius of the earth. From the law obtained by way of the generalisation of experimental data, none of this can be deduced.
Let us take as a second example the well-known Boyle–Mariotte law. It states that at constant temperature the pressure of a gas is proportional to its density, or inversely proportional to the volume of unit mass, that is:
if is the pressure and the volume of unit mass. This law too is depicted in experimental physics as a generalisation of systematically conducted analytical experiments and observations on various gases at various pressures, and so forth, and it was indeed obtained in this way.
In theoretical physics, however, this law is derived as a particular case from certain general representations and laws of motion of gas molecules. It is assumed that the gas consists of separate molecules moving with various velocities, whereby it is supposed that the velocities are distributed along the three coordinates according to the Gaussian curve of errors, and that the mean square of the velocities is proportional to the temperature and, consequently, is constant at constant temperature. Moreover, it is assumed that the general laws of motion are valid for molecules too. On the assumption of these general representations and laws, the Boyle–Mariotte law can be derived as a particular case.
Let us indeed denote the weight of the separate molecules of the given gas as equal to , the number of molecules moving with a velocity between and in unit mass as equal to ; then the number of molecules passing, in unit time and with this velocity, through unit surface will be equal to , and their quantity of motion ; if we take the mean square of the velocity of the corresponding normal component, then the pressure on the wall will be equal to one third of this magnitude, that is
if is the pressure and the mean square of the velocity. But since is nothing other than the weight of unit mass, that is, the density , we have
or
and this is the Boyle–Mariotte law
if we recall the general assumption that the mean square of the velocity of the molecules depends on the temperature and is consequently constant at constant temperature.
In this case we have again derived the law generalised from the data of experience as a particular case of more general laws and propositions. Again we see that in the second case the same law states more in its content: it contains statements also about the molecules and their motion, of which the law obtained by the causal-analytical path by means of generalisation says nothing, and which are not given immediately in experience. Again the question arises, whence we take these general representations and propositions, which, on the one hand, are abstract and general, and, on the other, have a richer content. From these general, abstract representations and propositions one can, as is well known, derive not only the Boyle–Mariotte law, but, on adopting the corresponding conditions, also the remaining gas laws obtained by way of generalisation from experience. We therefore speak of the kinetic theory of gases as a part of theoretical physics. These general representations and laws of the kinetic theory of gases contain the particular laws of motion of every gaseous matter, under whatever conditions they might manifest themselves, so long as the parts making up this matter—that is, the molecules themselves—are not subjected to changes; in other words, so long as no chemical changes arise in it. This leads us also to the answer to the question, whence and in what way these general abstract propositions and representations are obtained, as, for example, the Newtonian laws of motion in mechanics, the Newtonian general law of the attraction of masses, the propositions and representations of the kinetic theory of gases. They are not the result of the further generalisation of some empirical law, for example the law of fall, or the Boyle–Mariotte law, but the result of the bringing together into one of various empirical laws and particular data of experience, the condensation of all these data of experience and empirical laws into determinate general propositions and representations, which themselves already go beyond the bounds of immediate experience and are of necessity abstract, since they abstract the general, the essential, namely the general laws of motion proper to the given material state. Only on condition that we can abstract these general laws of motion, proper to the given material state, and represent the various particular empirical laws as a manifestation of these general laws of motion under determinate conditions—only then do we speak of a theoretical or general science of the given domain. The collecting and exposition of the particular laws of the given domain, obtained by way of generalisation, is the task of descriptive and experimental science.
What has been said is correct not only for mechanics and for other domains of physics. The very same we find in chemistry, where, on the basis of the same criterion, we distinguish general or theoretical chemistry from special, organic and inorganic chemistry. General or theoretical chemistry is occupied only with those processes and laws which are common to all chemical processes and valid for all of them, under whatever special conditions these processes might proceed. It expounds those laws of motion which are accompanied by a change in the structure of molecules, whatever special structure these molecules might possess. It formulates these laws as particular cases, under the given conditions, of certain more general representations and propositions about the structure of molecules and their motion; it derives them from these propositions. Thus, the laws of electrolysis found by Faraday by way of the generalisation of the results of causal-analytical experiments are, as is well known, derived in theoretical chemistry from general representations and propositions about the structure of molecules and about their electrolytic dissociation. Thus, there are derived in theoretical chemistry from general representations about motion and structure those regularities by which the various reactions proceed with respect to the equilibrium that is established, the velocity, and the course in time; and the laws of chemical dynamics thus obtained constitute the subject of theoretical chemistry. The exposition of the various laws of electrolysis, of chemical statics and dynamics, on the basis of the generalisation of the corresponding observations and experiments, would not yet be theoretical chemistry. Only when we can represent these laws on the basis of representations and general propositions obtained by way of abstraction—that is, of the condensation of various empirical propositions about the motion of chemical compounds—when we can derive them as particular cases, only then and only in so far do we speak of theoretical or general chemistry. The examples adduced above sufficiently elucidate this.
Here, consequently, our equation of the general with the theoretical, on the one hand, and of the particular with the descriptive-experimental, on the other, is correct.
How then does the matter stand with biology? At first glance it seems that between physics and chemistry, on the one hand, and biology, on the other, there is an essential difference. Physics is occupied with the laws of motion of non-living matter, in so far as no changes of chemical structure take place in it, and independently of the form in which these bodies exist, of whether the given motions actually occur in nature, and to what regularities, with respect to the existence, distribution and changes in time, the manifold concrete physical forms of motion are subject. Chemistry too is occupied with the laws of chemical structure and its changes, independently of where and in what quantity these various chemical bodies occur in nature, and to what regularities, with respect to the existence, dissemination and change in time, the manifold concrete chemical bodies are subject in nature. Thus, for example, the physics of gases is not occupied with the regularities of the changes of pressure, of the formation, and so forth, of the accumulations of gases existing in nature and of their changes in time. With this, partly, from practical motives, applied physics of gases is occupied: meteorology, which studies the regularities of the winds, and so forth. Hydrodynamics is not occupied with the regularities to which the liquids occurring in nature, with their motions, arising and changes in time, are subject; thus, for example, it is not occupied with the regularities of the motion of the rivers occurring on earth and of their changes in time. The same relates also to electrodynamics, and so forth. Chemistry too is not occupied with the regularities to which the existence, quantity, formation, dissemination and changes in time of the diverse chemical compounds in nature are subject. With this, partly, the division of applied chemistry in geology is occupied. Biology, on the contrary, has to do with the living beings existing in nature, with the natural bodies as they are given in nature, and its first task was, as we see, the investigation precisely of the existence of the form of these living beings, as well as of the regularities of their formation, distribution on earth and their changes in time.
However, this difference is only a difference in the historical development of the direction of investigations in these sciences and is not conditioned by the very essence of the subject.
We think that the difference in the development of the direction of investigations was conditioned by the different significance which inorganic and organic, living nature and their laws of motion have for mankind, for the satisfaction of its practical needs.
Inorganic nature and its laws of motion always had for man the significance that he could, under the given conditions of existence, make use of them and direct them, could with their aid fashion his first working tools, and subsequently build his machines. The first, primitive working tools were simple machines, such as the lever, and so forth. Mechanics developed out of the practical need to perfect these machines; out of the same needs there developed dynamics, in order to build better implements or to make better use of them with the aid of the laws of ballistics. Hydrodynamics arose out of the practical need to build hydraulic pumps and machines on the basis of the laws of liquids, but not out of the need to make use of the natural water-forces as such, since for the better use of the latter there are needed corresponding machines with a corresponding knowledge of the laws of hydrodynamics and mechanics. Aerodynamics developed especially in connection with aeronautics. Thermodynamics too developed in connection with the development of the steam engine, and not in connection with the question of where and in what form there occur in nature the transformations of energy useful to us, and so forth. The very same relates in great measure also to chemistry, which arose out of the practical need to create, from natural products, compounds and substances useful to man, so as not to be dependent on the products available in nature, which occur only in a limited quantity. The predecessor of modern scientific chemistry was alchemy, which gathered many valuable chemical observations and items of knowledge in the striving to create from worthless substances precious gold.
But organised living matter, living beings, always played for mankind a different role. Plants and animals were always for man conditions of existence of the first importance; they were his food, and indeed in the form in which they occur in nature; man could make use of them only in so far as he knew their distribution, conditions of existence in nature, reproduction and mode of life. The first means by which man procured food for himself were fishing, hunting, and the gathering of certain fruits occurring in nature. Even at the present time there can as yet be no question of the manufacture of food products independently of the living beings, plants and animals, occurring in nature. By this was conditioned the fact that man was in the first instance interested in the cognition of the various living beings occurring in nature, of their distribution and reproduction, of their mode of life and of those regularities to which the conditions of their appearance in nature are subject. By virtue of this, in biology the natural-historical moment developed first and most strongly, whereas the experimental direction, investigating the general laws of motion of living matter independently of the accidental conditions of its appearance, arose only later. In the inorganic sciences, however, we observe the reverse picture. It would, however, be incorrect to think that this difference is conditioned by the very object subject to investigation, by a difference in the state and laws of motion between living and non-living matter. Non-living nature has just as much its history as the living, and the ultimate aim of every science consists in this, that it should investigate the regularities of this history of nature and, on the basis of them, foresee phenomena and govern them.
But in order to find the regularities of the history of living or non-living nature, it is by no means sufficient to know the conditions of existence, distribution and change in time of the given diverse forms of phenomenon and motion. Only in that case can we speak of a historical regularity, if from the laws of motion proper to the given matter we can deduce the necessity, the law-governedness of these changes in time, of the arising and disappearance of some forms of motion and the appearance of others—if, on the basis of these general internal laws of motion, characteristic of the given matter and proper to it alone, we can show that these regularities and their changes in time are accomplished, with internal necessity, through the particular and the accidental. There exists only one domain of knowledge, one science, where this task has been resolved, where, on the basis of general internal laws of motion obtained by way of far-reaching abstraction—that is, of the condensation of the most diverse empirical propositions—the necessity of historical regularity in the above-indicated sense has been shown. This is the Marxist doctrine of society. The general laws of motion of human society discovered by Marx gave a method for the investigation of any form of society, and precisely for this reason, on the basis of them, one can, by means of the analysis of any social order, predict with the inevitability of a law of nature the changes (and their direction) of this order, and also act in a planned manner upon the course of these changes.
Bringing together all that has been said, we see that the difference between the inorganic sciences and biology is conditioned only by the historical development of these sciences, and not by fundamental differences of the aim which they set themselves, or by differences in the method with the aid of which this aim is or can be attained.
Thus, our conclusions regarding the theoretical sciences are fully applicable to biology too. Moreover, we see in the inorganic sciences too that the historical regularities of non-living matter, the inevitability of the arising, ageing, perishing and changes of the heavenly bodies, as well as the direction of these changes, in so far as they have hitherto been investigated with success, could be investigated precisely only on the basis of, and with the aid of, the general representations and laws of motion obtained by theoretical physics and chemistry. Without the general laws of motion of mechanics, of the force of attraction, the laws of gases, of thermodynamics, the laws of radioactivity, and so forth, the natural sciences would not have arrived at the modern representations concerning the arising and history of the heavenly bodies. The very same relates also to the history of our earth, of its strata, and so forth.
We now come to the second point of our scheme standing in need of explanation—this is the question of the content and significance of evolutionary theory and of its relation to the general theory of living matter within the framework of theoretical or general biology. As concerns the content of evolutionary theory, it is undoubtedly correct to represent the latter as an achievement of theoretical biology in the sense indicated above. Evolutionary theory is not merely a generalisation of immediate empirical data or of an empirical law. It is a condensate of a great number of various empirical data and regularities of comparative morphology, descriptive biology, and so forth, and goes beyond the bounds of immediate experience; it states a general regularity, obtained by way of abstraction and on the basis of certain general representations about living beings—a regularity stating that living beings on earth, animals and plants, arose one from another in the course of the history of the earth, that they possess a common genealogical tree, that is, stand in a more or less close “blood” kinship with one another. All the particular empirical data obtained by comparative morphology, zoogeography, palaeontology, and so forth, can be represented as certain particular cases of this general principle, or of this general regularity. Even more than that: evolutionary theory possesses also a historical-temporal moment, in so far as it contains within itself the necessity of the appearance of new forms and the perishing of others. But it states no historical regularity. From evolutionary theory in its modern form and with its modern foundation, nothing can be deduced concerning which forms of living beings could or had to appear under determinate conditions and in a determinate historical period, which regularities are accomplished with necessity in the course of the history of living beings through the particular and the accidental. Nothing can be said about wherein and in what direction the later forms of life had to differ from the earlier. The principles of evolution and of evolutionary theory in their modern form contain the proposition that the various forms of living beings, that living matter, have their history, but about the laws of this history they say nothing and permit no statements. That evolutionary theory, as Darwin founded and elaborated it, did not go beyond these bounds, is of course no accident. This stands in connection with the fact that it is a theoretical achievement precisely of the first, descriptive phase of biology. According to the then degree of development of science, it could not go further. The deficiencies of its epoch were reflected in it. True historical regularities in the sense expounded above can be recognised only on the basis of the general laws of motion proper to living matter. Only by knowing these general laws of motion can we show how they change their form upon a change of conditions, and in what direction such a change is possible and must set in. Upon what general laws, characteristic of living beings, could Darwin then rely? Upon those which could be obtained by way of generalisation from the empirical data of the first, descriptive phase of biology—that is, upon the laws of reproduction, variability and heredity. In the foundation of his doctrine Darwin made full use of them. But he could rely only on these general laws of the manifestations of living matter, and not on the internal laws of motion of this matter itself. A general theory of living matter then did not yet exist. Neither were there special laws of motion of particular phenomena. There was no theory of heredity in the sense of modern genetics, which attempts to derive the phenomena of heredity from the laws of motion proper to the hereditary substance. A similar theory of variability, reproduction, and so forth, is lacking to this very day. It is therefore understandable that in modern biology there are observed attempts on the part of genetics to deepen evolutionary theory, to reduce it to a theory of heredity. That these attempts remain unsatisfactory is explained precisely by the fact that genetics itself can operate only with those special laws of motion of living matter on which the laws of heredity are based, and not with those universal laws of living matter which are obtained by way of condensation—that is, of abstraction—not only from the phenomena of heredity, but also from the phenomena of growth, reproduction, adaptation, development, and so forth—that is, from the empirical data also of physiology, the mechanics of development, and so forth. Only on the basis of, and with the aid of, such general laws of living matter can a successful further deepening of evolutionary theory be attained; only on the basis of, and with the aid of, such laws—that is, of a general theory of living matter—can evolutionary theory be elaborated and deepened into a true historical theory which embraces the historical regularities and their necessity and permits statements regarding them.
In its modern form, evolutionary theory not only permits no statements regarding historical regularities—that is, regularities concerning the various species of animals and plants in comparison with their past; it even fundamentally denies any regularity in the sense of a regularity accomplished in time with necessity through the particular and the accidental—that is, in the sense of directedness in time. Thus, we have in modern evolutionary theory a paradoxical position, as it were, namely a theory which postulates and founds the history of living beings, but fundamentally denies the existence of regularities of this history. In it there reigns the moment of the accidental, covering over the moment of the historically necessary, the law-governed.
This paradoxical phenomenon has two causes: in the first place, any science must categorically reject every theory which assumes that historical regularities, in the sense of directedness, of orthogenesis, are called forth by immanent, immaterial, directing forces, or attempts to explain these regularities in such a way—that is, in a vitalistic direction. Therefore one ought, as a matter of principle, to reject as unscientific all explanations and theories proceeding from such assumptions and leading, or even merely capable of leading, to them. In the struggle against these tendencies, however, scientific biology developed, which, adhering to the postulate of the history of living beings, ever more proved the presence in this history of the element of the accidental.
In the second place, the moment of the historically necessary, the law-governed, accomplished through the particular and the accidental, could not and cannot be recognised, grasped and materialistically explained, if it does not rest upon the general laws of motion proper to living matter. These two causes are combined in modern biology in such a way that, in view of the absence of a materialistic explanation for historical regularities—that is, for those which are accomplished with necessity in time in a determinate direction—biologists fundamentally deny these regularities and their existence, representing them as contradicting the materialist-dialectical, scientific mode of thought and world-outlook.
But that such a point of view is incorrect is shown to us by the fact that, at a determinate degree of development of the productive forces, the direction of development of the social order is predetermined with necessity and “the wheel of history cannot be turned back”. It is sufficient, for example, to know Lenin’s argumentation against the Narodniks, based on the general laws of motion of society, in order to see that the assumption of a historical regularity in the sense of one which is accomplished with necessity in a determinate direction in no way contradicts dialectical materialism, but, on the contrary, constitutes an essential element of the latter. On the other hand, by the example both of Marxism and of certain of the above-adduced domains of the inorganic natural sciences, we see that to grasp and explain these historical regularities materialistically—that is, scientifically—is possible only on the basis of, and with the aid of, the general laws of motion of the given matter.
By this we have given an answer also to the question of the interrelations between the theory of evolution and the general theory of living matter within the framework of theoretical biology according to our above-adduced scheme. The theory of evolution stands in need of deepening in order to become a true historical theory of living matter. This further development can be attained only on the basis of the general laws of motion of living matter. Therefore the immediate task of theoretical biology is the elaboration of the general laws of motion of living matter, that is, of the theory of the latter. The task standing before theoretical biology, to the resolution of which it is approaching, consequently consists in this, that it should represent and deduce the empirical laws and data of descriptive and experimental biology with its divisions as particular moments of the development of more general laws and representations about living matter which go beyond the bounds of immediate experience. These general laws must be laws of motion inherent in living matter, that is, proper to it alone. But they must constitute the laws of motion proper to it everywhere and always, the form of its existence, in whatever special kinds they might manifest themselves. Therefore they must be valid for every living matter and for it alone. They must permit us to deduce, from the analysis of concrete conditions, the appearance of special forms of motion and the direction of their change. This is a long path. The first and greatest step in this direction was made by the evolutionary theory founded by Darwin. The further considerable preliminary work was accomplished by the developing special, experimental sciences, which approached the investigation of those laws of motion of living matter from which the particular regularities of the manifestations of this matter can be deduced—in the first place the doctrine of heredity, or genetics, which strives to represent its regularities as particular cases of more general representations. The following step must be made in the direction of a general theory of living matter, in order, with its aid, to approach the resolution of the tasks indicated above. To elucidate, in so far as this is already possible at present, to show that this path is open to us, and to make a further step in this direction—this is the aim of the present book.
Chapter 1: The Principle of Stable Non-Equilibrium
On the basis of certain phenomena, we designate some material systems as living beings, as living systems [живые системы — zhivye sistemy]. The aggregate of those phenomena on the basis of which we designate a system as living, or the internal conditions necessary for this aggregate of phenomena, we call life. In ordinary colloquial speech one says, for example: “The animal still shows signs of life,” and by this is meant that the conditions for the manifestation of the said phenomena in the animal have still been preserved. But if one wishes to approach a definition of these phenomena, or of the conditions with which they are bound up, one runs into a striking and seemingly contradictory difficulty. On the one hand, we wish to characterise certain material systems which, on the basis of certain properties, we designate as living, and thereby to mark them off from the remaining systems; on the other hand, we find it hard to indicate even a single property that would be characteristic of these systems and would distinguish them from the rest. The contrast between the clarity and assurance with which the designation “living” is employed in both ordinary and scientific language, and the difficulty and indeterminacy, as well as the uncertainty,The source reads «достоверностью» (“certainty”), evidently an OCR corruption of «недостоверностью»; rendered here by sense. with which science attempts to give a definite and clear sense to this designation, is truly striking. The easiest way to make this contradiction clear to oneself is perhaps to take the textbooks of biology and to establish that in their introductory and general sections they speak of the difficulty and even the impossibility of any such demarcation, whereas in the description of a living being and the interpretation of vital phenomena no doubt ever arises in the text as to whether the object under consideration, or the phenomenon under consideration, is to be reckoned among living beings or vital phenomena, and whether, consequently, they rightfully occupy a place in the textbooks of biology.
This state of affairs has led, in the concrete exposition of biology, to a tacit agreement that, in expounding biology and in marking it off as such from the other sciences, it is best to rely upon this instinctive-empirical demarcation; and therefore the description and analysis of vital phenomena are carried out neither from the standpoint of the general characteristic properties and regularities that govern all vital phenomena (living systems), nor from the standpoint of those conditions which are necessary for their manifestation.
But here we shall not adhere to this custom; rather we shall attempt to single out those properties and laws that are characteristic of living systems, and shall see in what manner these properties and laws are manifested in various living beings.
There have, of course, been not a few attempts to single out these properties and to give a definition of life, but these attempts have for the most part had the defect that their initiators did not take them seriously enough, in that they did not attempt to verify the correctness of their definitions against vital phenomena, since this was done in too general a form, so that it was impossible to confirm or refute them by means of concrete observations or experiments, that is, to verify them at all.
We shall in our turn attempt to give such a definition of life as could be verified for correctness, and shall endeavour to carry out this verification.
Our task consists in combining and expressing in the form of one or several laws that which is proper to all systems that we designate as living, and which is characteristic of them alone.
Change of state of living systems under constant external conditions.
Proper to all living beings is, first of all, a spontaneous change of their state, that is, a change of state which is not caused by external causes lying outside the living being.The source omits the negation here («изменение состояния, которое вызвано…»); restored by sense, as required by the definition of spontaneity. No one will define a body or a material system as living if he can detect in it no changes at all, or can detect only such changes as bear a purely “passive” character, that is, which are undoubtedly the result of changes of the environment or of the influence of external forces, without any “active” participation of the system itself. Thus, if a material system is in complete repose and moves, for example, only when and whither it is moved or thrown, and only in accordance with its mass, inertia, and frictional resistance; if it has the temperature of the surrounding environment and is heated or cooled only when and to the extent that the temperature of the surrounding environment changes, and indeed in accordance with its heat capacity; if in the material system no chemical reactions occur, or they occur only when substances capable of reaction are introduced into it from without—and only to such a degree and at such a rate as correspond to the law of mass action and to chemical kinetics under the given initial conditions in the material system, even were these reactions very strong or conditioned by the presence of various ferments, and so forth—then no one will think of defining this material system as living, that is, no one will see in it a living being. Consequently, one of the requirements that we set for a material system, in order to designate it as a living being, is not fulfilled in the cited examples. In what does this requirement consist, and in what manner can it be more precisely formulated?
If under the existing conditions, without change of the surrounding environment, the system is not to be in complete repose, if changes can occur in it “spontaneously,” then this means that there must be in the system such accumulations of energy as, under the conditions prevailing in the system and under unchanging conditions of the surrounding environment, can be discharged. There must exist in the system such differences of potential as, apart from external assistance—that is, without change of the surrounding environment, or, what is the same, without external action upon the system—can be discharged, that is, equalised, whereby the energy liberated in this equalisation, in this discharge, may manifest itself in various ways, be it as mechanical work or as the formation of heat, of electrical energy, and so forth. In what the energy thus liberated must manifest itself, in what work—in this respect we set, for the time being, no requirements.
This first requirement, which we unconditionally make of a material system in order to designate it as living, is of course not sufficient, since it can be fulfilled by any “wound-up” machine that has been set going. A wound clock with a pendulum shows changes independently; it runs without external assistance, without change of the surrounding environment, while the potential energy of the wound weights is gradually equalised, that is, the weight descends and is thereby in part converted into the kinetic energy of the motion of the pendulum, of the motion of the hands, and so forth; a wound pocket-watch has potential energy in the bent spring of the watch, in the form of so-called energy of deformation, which is gradually equalised and can be equalised without external assistance, whereby this energy of deformation diminishes and reveals itself in the form of the work of the gear-train and the motion of the hands. A heated steam-engine has potential energy in the form of the difference of temperature that exists between the heated boiler and the condenser; this difference of temperature is equalised and can be equalised without external assistance, and then manifests itself in part in the form of the various mechanical works for which the machine is constructed. A charged accumulator shows at its poles a difference of electrical potential which, in the case where the two poles are joined by a conductor, is equalised and can be equalised without external action, whereby the electrical energy can, depending on the construction of the conductor joining the system, be given the form of various energies, and so forth.
We see that the requirement that a system should, spontaneously, without external assistance—consequently, without change of the surrounding environment—show changes and perform various works, or, what is equivalent, that it should possess potential energy which is discharged, should possess differences of potential which can be equalised under the given conditions without external assistance—that this requirement can be fulfilled by any “wound-up,” “set-going,” “charged” machine.
These machines that have been set going, however, we shall never call living. And this means that of those material systems which we designate as living, we make yet further certain requirements. However, many natural scientists and biologists have a tendency to regard these other requirements, which we connect with the designation “living,” as unfounded, or at least as requirements of a non-fundamental character, in that they say that living systems—consequently those systems for which we retain in ordinary language the designation “living,” “living organisms,” or “living beings”—for the most part represent nothing other than machines that have been set going, whose operations proceed, like the operations of all other machines, according to the laws of physics and chemistry and the conditions prevailing in the system and in the surrounding environment. Not doubting for a moment that the physical and chemical processes proceeding in the systems designated as living are subject to the laws of physics and chemistry and are clearly determined by them, as well as by the conditions of the system and of the surrounding environment, we nonetheless consider that our task naturally consists not in simply resting content with this, but in seeing whether these other requirements, with which we connect the designation “living,” cannot in fact be established and in what they properly consist—that is, whether these material systems do not, among others, after all exhibit special properties or regularities which are proper to them and distinguish them from the remaining systems, and which modify the laws of physics and chemistry.
Change of state under change of external conditions
Above we said that no one will designate a body or a material system as living when he can detect in it no changes at all, or only such as bear a purely “passive” character, that is, which are undoubtedly the result of a change of state or of the influence of external forces, without any special “active” assistance of the system itself.
If our first requirement amounted to this, that a living system shows a change even without external assistance—consequently, with the surrounding environment unchanged—is capable of work, and consequently possesses potential energy and can convert it, then the expression “or only such as bear a purely ‘passive’ character,” and so forth, contains within it yet another requirement, which relates to the behaviour, that is, to the processes in the system under changes in the conditions of the surrounding environment. As we shall now see, this second requirement presupposes, as a necessary condition, the fulfilment of the first requirement.
To make clear to ourselves the meaning of the second requirement, we shall again return to the previously cited examples. If we cause a force (P) to act upon a material system with mass (m), then the body receives an acceleration (a), the magnitude of which is established by the relation:
Force = mass × acceleration.
When to this is added friction as well, then, plainly, the force must overcome it too, and a greater force will be required in order to produce the same acceleration, that is, the relation will be obtained:
Force = mass × acceleration + friction.
Further, the body can, by means of certain devices, be made to move along a determinate path or along a determinate surface, for example along a circle, when it is tied to one end of a thread the other end of which is fixed at one point: if the body revolves about this point with a certain velocity, it will always be constrained, owing to the thread, to move in a circle; or, for example, when the body lies on an inclined plane, and so forth. The action of such conditions, which constrain the body to remain on a determinate path or surface, we may replace by forces that produce the same action, and shall designate these forces as “constraint forces,” in distinction from the “driving forces” acting upon the body. The body of mass (m) experiences, under the action of these constraint and driving forces, an actual acceleration (a). In mechanics there exists a general law of motion, called d’Alembert’s principle, according to which the sum of the constraint and driving forces and the negatively taken product of the magnitude m × a (mass multiplied by the actual acceleration), which is also designated as the force of inertia, is equal to zero, that is, the constraint forces, the driving forces, and the forces of inertia are always in equilibrium:
where Z denotes the constraint forces, P the driving forces.
After these brief explanations let us now attempt to consider what is to be understood by saying that a body, under the action of a driving force, moves only in accordance with its force of inertia, its mass, or its frictional resistance, and what we require of living systems, namely that they should move not only in accordance with this.
Let us begin with the very simplest example: upon a body lying on a horizontal surface there acts a driving force in the direction of the surface, that is, the force displaces the body along the surface.
In this case the driving forces are represented by: the acting force of traction (K) and the force of gravitation, which the earth produces, mg (g being the acceleration of gravity); the constraint force in the given case is equal in magnitude to the force of gravity and directed in the opposite direction, since it is precisely this that does not allow gravity to manifest itself, and is consequently equal to mg; and if the body experiences an acceleration a, then the above-mentioned law of motion states:
that is,
In order to bring the body out of the state of repose and at the same time impart to it an acceleration a, the force must be equal in magnitude to the inertial resistance of the body and opposite to it in direction. If friction is also taken into account here, then the force must overcome it too, that is,
and when the body has already been set in motion and is, by means of the force, to be advanced further at a uniform velocity, then, since the acceleration—and consequently the force of inertia too—will be equal to zero, the driving force will have only to overcome friction, that is, be equal to it:
But the force needed to displace a living animal almost never corresponds to the force of inertia and to friction, save only when the animal is dead or behaves quite “passively” with respect to the force of traction or the force of impulse acting upon it; the animal “reacts” to traction or impulse by running away or by producing resistance, that is, it exhibits a resistance which will be greater or less than what corresponds to the force of inertia or to friction. The point, plainly, is not that d’Alembert’s principle of mechanics is invalid, but simply that, besides the external constraint and driving forces acting upon the system, the system develops during the traction or impulse certain internal forces which we have here not taken into account, that is, the motion occurs not only in accordance with the force of inertia and friction, but also not without “active” assistance of the system itself.
In exactly the same way it can be shown that in animals living in water the effect of an impulse is for the most part conditioned not only by friction against the water, by the force of gravity, by the moment of inertia, and by the applied force in accordance solely with the laws of motion of mechanics, but that to these are added still further forces which the living system develops during the action of the external force and by this changes the motor effect. In what direction the system must develop this force, and in what manner it must change the motion—of this the second requirement as yet says nothing either.
Considering further the action of mechanical forces, we find that what has been said remains valid not only for the motion of a system under the action of a force, but also, for example, for the changes of form which a system undergoes under the action of traction or pressure; these changes of form are determined, in living systems, not only by pressure or traction, or by the modulus of elasticity of the tissue which they possessed prior to the given action: the protoplasm too influences this action of deformation, in that it develops during the pressure or traction certain forces which lead to a change of the modulus of elasticity, and so forth. This second requirement, which amounts to this, that living systems must exhibit not only those changes which are an evident result of external changes without any “active” participation on the part of the system itself, must be applied in quite the same manner to other, non-mechanical actions as well: it is known that a body, upon the heating of the surrounding environment, itself becomes heated, whereby this rise of temperature is determined by the quantity of heat imparted and by the heat capacity of the various parts of the material system; but in living systems, as we know, the rise of temperature is not determined by this alone, since the living system “reacts” to the influx of heat by means of a change of heat-production and heat-emission. Likewise, an exact knowledge of the concentration of the substances present in the cell or tissue and entering into reaction, of the reaction constant, and so forth, is not sufficient to determine, according to the laws of chemical kinetics, the rate of processes in living systems and the final state to which they lead, since these systems develop during the reaction forces that change the conditions and the course of the chemical reaction.
This “active” counteraction to the external forces acting upon it and to the changes of state of the surrounding environment, which we require of a material system in order to designate it as living, has, as we have seen from the examples, as a necessary presupposition the fact that in the system itself, during external changes of state, there occur such changes as are not merely a consequence of the changes of state of the surrounding environment and of the initial conditions existing at the action of the forces. For this, however, the first requirement must be fulfilled, namely that the system should be able to exhibit a change of state “spontaneously,” that it should possess potential energy which it can employ for the development of force here necessary during the external action. But the second requirement contains, of course, more than the first—namely, that the existing differences of potential should be equalised in a certain dependence upon the external actions, or that the energies liberated upon their equalisation should be applied to the performance of such works as stand in a certain dependence upon the changes of state of the surrounding environment, whereby they influence the effect of these latter, that is, during the external action they so change the conditions existing in the system that thereby a change of action is conditioned. In what direction this work must occur and what change must ensue—of this the second requirement says nothing either: it requires only that in living systems, during external actions, such work should occur as changes the initial conditions of the system, and consequently the effect of the action of the surrounding environment.
If we now ask ourselves: is this second requirement, taken together with the first, sufficient in order to say that those systems in which they are fulfilled will be designated as “living,” then we must answer that this is not sufficient, and above all for the reason that it does not represent an unconditional requirement. We do not require that living systems should unconditionally—that is, in every case and to every external action—react with work that changes the effect of the action, but that they should show not only “passive” changes, which can be determined unequivocally on the basis of the external action upon the system and from its initial conditions. If, however, this does not represent an unconditional requirement, then one can nonetheless picture to oneself such constructions as respond to certain external actions by the development of forces influencing this action. For this, these systems must possess certain mechanisms of release which, under certain external influences, come into action and produce in the system certain processes yielding energy, whereby the liberated energy is applied to influencing the effect of the external action. As an example there may serve a cart set in motion by an accumulator, with a mechanism which, upon an impulse, switches in a higher voltage and thereby, during the impulse, imparts to the cart a higher kinetic energy, thus influencing the effect of the impulse, which will be other than that calculated from the force of the impulse at the very moment of it, from the velocity and mass of the cart, and so forth. A second example: an elastic spring with a source of electric current and a mechanism which, upon the stretching of the spring, switches in the current and passes it through the spring, whereby the latter is heated and changes its modulus of elasticity. Or, for example, a mixture of substances capable of reaction, joined with a thermoelectric element which, upon being heated, sends a current through the mixture and calls forth an exothermic reaction that would not have come about without the current at this temperature; in consequence of this the heating will be greater than that calculated from the quantity of heat applied for the warming of the element, the reaction constant, and so forth.
All these constructions are an example of systems that satisfy the second requirement, inasmuch as they possess devices by means of which they change, upon an external action (impulse, traction, supply of heat), through processes yielding energy at the expense of the differences of potential existing in the system, the influence of this external action.
In these examples we see that our second requirement nonetheless contains considerably more than the first. Whereas a cart set in motion by means of an accumulator, alone and without the mentioned mechanism, satisfies the first requirement because processes occur in it “spontaneously,” without external assistance, it does not satisfy the second requirement without the mentioned mechanism, since, although the motion of this latter also influences the action of an impulse upon the moving cart, the effect can be calculated from the velocity, the mass of the cart, and the force of the impulse, whereas the mentioned mechanism leads to the system’s performing work that changes the influence calculated from these magnitudes. A mixture of substances capable of reaction, in which the reaction is still proceeding, also satisfies the first requirement, because processes occur in it without external action, consequently also without the supply of heat; but the result of a certain rise of temperature can be calculated from the quantity of heat supplied and the reaction constant, in the case where the mentioned mechanism does not exist: this latter is necessary for the fulfilment of the second requirement, since only by means of it does the supply of heat condition not merely a “purely passive” change of the chemical equilibrium in accordance with the raised temperature, but also a further, energy-yielding process that changes the direction or degree of the change of equilibrium. To avoid misunderstandings, it should here be added that, given the existence of the said mechanisms, the change of state called forth is naturally determined unequivocally by the initial conditions in the system and by the external actions, since the mentioned external actions necessarily call forth the activity of the said mechanisms; taking into account all these conditions, the change of effect can be determined unequivocally.
If, then, the second requirement is fulfilled only by means of complex constructions, machines provided with certain devices, then these complex constructions too are not yet designated by anyone as “living.”
From this it clearly follows that, to the designation “living,” besides the fulfilment of the first and second requirements, we attach the fulfilment of yet other conditions. It should nonetheless be noted that, whereas the constructions which we cited as examples fulfil the second requirement only for a quite determinate and only single external action, this second requirement, in the systems designated by us as living, is very widespread and is fulfilled at almost every external action and at any repeated actions. In order to build systems that would correspond to the second requirement at almost any, and moreover repeated, external actions, one would have to devise extremely complex mechanisms.
Therefore, the fulfilment of the second requirement is often regarded as an essential mark of a living system and is designated as “irritability” [раздражимость — razdrazhimost′] or “excitability” [возбудимость — vozbudimost′].
Irritability, excitability.
By “irritability” or “excitability” is in general designated that property of living organisms by virtue of which they respond to changes of state of the surrounding environment, that is, to external actions, by changes of state, that is, by processes which are difficult to derive from the external action of the so-called stimulation; the action does not correspond to the force of the external action—insignificant actions are released by intensive processes: very often there is also lacking a correspondence in place: the “stimulation” acts at one place, but the process conditioned by it occurs at quite another place in the organism. Consequently, the concepts of “irritability” and “excitability” belong to those concepts which were created in order to single out and characterise the special behaviour of living systems with respect to external actions.
Let us now see in what this special behaviour properly consists, what is meant by it, and in what relation it stands to our first and second requirements.
If the matter consists only in the quantitative non-correspondence between the external action and the change of state called forth by it, between the “stimulation” and the response reaction, then this means that here we are dealing with so-called phenomena of discharge. There exists in the system potential energy in the form of various differences of potential, which can neither be discharged nor equalised without external assistance; an insignificant action from without is sufficient, however, to produce this equalisation, whereupon, then, quite independently of the discharging action, enormous quantities of energy are liberated and a very strong action may be attained.
Similar phenomena of discharge are, of course, known in non-living systems as well, for example in strong explosions, which are called forth by means of a light pressure on a button, whereby an electrical connection is made that transmits an electric spark or current through the explosive mixture; here too the place of action of the external change and of the response process are different. If, in this property of living systems which we designate as irritability or excitability, the matter really concerned only that property, that in living systems such phenomena of discharge are very often encountered, then there would be little sense in singling out this property in particular, and it would be difficult to understand why, nonetheless, this is always done.
Many biologists arrive at the conclusion that to single out the phenomena of stimulation into a special group of phenomena, which would be in a certain respect characteristic of living beings, is correct only in so far as our imperfect knowledge does not yet afford the possibility of analysing them.
Hartmann, for example, writes: “If it were possible by contemporary means of investigation fully to cognise and analyse the chemico-energetic processes occurring in the so-called phenomena of stimulation, then there would be no need of a special delimitation of the concepts of this group of vital phenomena, which are united under the concept ‘phenomena of stimulation.’”
“But since at present we are still very far removed from this goal, such a delimitation and separate consideration of the phenomena of stimulation are necessary in the study of the organic process. This is especially necessary when the physiological change of the stationary processes in the system as a whole that serves as the stimulation is released by an action which, first of all, cannot be brought into any noticeable chemico-energetic relation with the stimulation and for the most part shows a striking disproportion between the magnitude of the stimulation and its action. In consequence of this, the phenomena of stimulation for the most part bear the character of so-called releases…”
”…It is known, however, that such processes of release are not confined to vital phenomena alone, but are encountered in inorganic systems as well, the difference consisting only in that in the latter the energetic relation can for the most part be clearly indicated, whereas in the former the cause and the response action for the most part diverge, and the physico-chemical processes occurring between them remain unknown.”[Author’s note:] Hartmann, M., Allgemeine Biologie, Jena, p. 16, 1925.
And in another place he writes: “…the so-called irritability represents, however, a phenomenon which, strictly speaking, can be characterised and defined only negatively, namely as a physiological action whose individual reaction chains in the organism itself are for the most part unknown.”[Author’s note:] Ibid., p. 629.
We nonetheless think that the concepts “irritability” and “excitability” were introduced with a special aim, since with the phenomena of stimulation there were bound up certain conceptions which contain not only this simple fact of disproportion and of ignorance of the intermediate processes. And even if, in the phenomena of stimulation, we do not know the whole reaction chain, a knowledge of the final effects is already sufficient to establish that, in the irritability of organisms, the matter concerns not only a negative mark, but also processes and conditions which are necessary in order to designate a system as living.
First of all, it should be pointed out that the said phenomena of release in non-living systems can, by means of a force corresponding to the actions, be called forth only once. The matter always consists in this, that certain differences of potential, certain tensions, cannot be equalised under the given conditions; but when the equalisation begins in consequence of an external action, then it proceeds to the end, and a second similar action will not be effective. A spring stretched by means of a thread will shorten strongly when the thread is cut and will assume its former immobile position, but a second cutting or slackening will produce nothing more. An explosive reaction mixture will explode with greater force from a light pressure on a contact and will attain the corresponding equilibrium of the reaction; a second pressing on the button, even after a long time, will produce no action whatever… By “irritability” or “excitability” is meant precisely that property of living systems, that they generally always respond in a certain manner to a stimulation, that they do not lose their “irritability” as a result of a single response reaction. When a muscle is brought into irritation by means of an electric current, it contracts; if it is brought into irritation a second time—it again contracts; and when the muscle, after too frequent stimulation, no longer responds to it, we do not say that it has finally lost the capacity for irritation, but that the muscle has become “tired”; consequently we know that after a certain interval the same stimulation will call forth the same contraction, and irritability will thus appear anew. This peculiarity of living systems is naturally bound up with the concept of irritability as a constant property of theirs. If, then, the mechanism of the individual response stimulations consists in fact in principle of a discharge of energy, just as, for example, in the slackening of a stretched spring or the explosion of a mixture, then with the concept of irritability there is also bound up the fact that, besides these discharges, in living systems there for the most part follows a new charge as well; and it is precisely this latter regularity of phenomena in living systems, and by no means the ignorance of the reaction chain, that has led to the establishment of the concept of irritability.
If we now see which of the requirements established by us must be fulfilled for irritability formulated in this manner, then it becomes clear that the existence of individual response reactions in the sense of processes of release is contained in none of our requirements—for the first requirement consists in this, that in living systems there must exist accumulated stores of energy, differences of potential, which “spontaneously,” consequently without external assistance, that is, with the surrounding environment unchanged, can be equalised and actually are equalised. We require, in other words, that the living system, under unchanging external conditions, should show changes—consequently that it should not be in repose, in equilibrium. A spring stretched by a thread or an explosive mixture in the examples we cited above remain, under unchanged external conditions, in repose; the spring and the mixture are in equilibrium under the given conditions until the external conditions change—until the spring is cut or the explosive mixture heated by a current passed through it. The circumstance which is often regarded as very characteristic of the phenomena of stimulation, namely that insignificant changes of external conditions lead to intensive processes, shows only that the system is in so-called unstable equilibrium, but this is not required of living systems; on the contrary, the first requirement contains the opposite condition, namely that the system should not be in equilibrium (whether stable or unstable) and that the existing differences of potential should be equalised “spontaneously,” precisely without any external assistance.
If, however, by irritability we mean precisely that property of living systems, that after a single response stimulation the irritability does not vanish, but continues to remain, as a characteristic property—that consequently, after the discharges, after the equalisations of the differences of potential, there regularly occurs a new charge, a restoration of the differences of potential—then this property contains within it our first requirement, since in order to produce this new charging, this restoration of the differences of potential, work is necessary, energy is necessary, which can be supplied by the system only in the case where in it “spontaneously,” without external assistance, processes occur that yield energy, that is, when our first requirement is fulfilled. In order to stretch anew a slackened spring, work must be performed; this work we perform by stretching the spring anew to its initial position, which it had prior to its slackening; here, consequently, work must be performed from without. In a muscle this work is performed after its contraction in consequence of stimulation, by means of processes that yield energy, whereby these processes occur in the muscle without external assistance. It is precisely by this change, proceeding usually regularly after each stimulation, supplying energy and anew restoring the differences of potential, that the muscle differs from a simple spring. This is also the reason why to a stretched spring we do not apply that concept of “irritability” which we apply to a muscle. Consequently, the fulfilment of our first requirement is a necessary condition for the manifestation of “irritability.” The second requirement is not contained in it. This latter meant that the equalisation of the differences of potential called forth by a release proceeds not as one would expect from the initial conditions, the rate of reaction, and so forth, but that it will be changed in its course by means of some energy-yielding process in the organism. This second requirement is, properly speaking, advanced, for example, by Mangold[Author’s note:] Mangold, Ergebnisse d. Physiol., vol. 21, 1923, cited after Broemser in Handb. norm. pathol. Physiol., vol. I, p. 278 ff. for the concept “excitation,” which he opposes to the concept “stimulation,” saying: “Excitation is every active change of the processes proceeding in a living organism,” whereas he defines stimulation as “every external change which is capable of so acting upon a living being that it, in the course of its vital processes, itself reacts by a change.” Correspondingly, Mangold draws a distinction between the conduction of stimulation and the conduction of excitation. As conduction of stimulation he designates such processes in which “the transmission of an external physical or chemical change occurs through the parts of the living organism, without active participation of the organism itself.” As conduction of excitation, on the contrary, he designates such processes in which “a physico-chemical change of an unknown kind is conducted further with the active participation of the living being.” According to this definition given by Mangold, the concept “excitability” thus corresponds exactly to our second requirement. Namely, excitability according to these definitions designates that property of living systems, that they can influence the course of a process called forth by means of an external physico-chemical action through active participation, consequently by means of some process yielding energy, requiring an expenditure of work on the part of the organism. This means that the course of a process called forth by means of an external excitation cannot be determined from the magnitude of this action and from the initial conditions of those parts of the system upon which the action occurs, since it is precisely these conditions that are changed by means of processes accompanied by an expenditure of work in the organism. In what direction this change of the process must occur, of this nothing is here said either; this active change may consist either in an acceleration or in an inhibition. With such a definition of excitation and stimulation, the fulfilment of our second requirement is not
a necessary, indispensable condition for living systems as a general one. Consequently, as with our second requirement, it is not obligatory that every external action should unconditionally be accompanied by an active change of the course, requiring an expenditure of work, expected on the basis of the initial conditions, that is, that it should respond with excitation. Only one thing is required: there must occur not only passive changes, which are determined solely by the action and the initial conditions. According to this, not every external action will be designated as a stimulation, but only such as are accompanied by excitation. One and the same external action will represent a stimulation or will not, depending on whether it calls forth excitation or not, according as the effect of the external action manifests itself in one way or another, as it would be determined from this latter and from the initial conditions of the parts of the system upon which the action occurred. With this restriction of the designation “stimulation” to only such external changes as call forth excitation, it is clear that the concepts of irritability and excitability are not separable from one another. On the other hand, the concept of irritability, as we established it above, namely as the property of living systems of responding to processes of equalisation called forth by external actions with other processes that supply energy and lead to a new raising of the equalised difference of potentials—this concept (if we refer to those processes called forth by external actions also the more or less distant from one another, but regularly occurring, processes) coincides fully with the concept of excitability and contains within it our second requirement as well, since we do not obtain, as the effect of the action, the expected disappearance of the differences of potential. Thus, although stimulation and excitation are different, they are nonetheless transformed under certain conditions into one another.
Excitability or irritability defined in this manner—which expressions we now employ in their real connection in the above-mentioned sense, as equivalent—contains, consequently, within it our second requirement as well, the fulfilment of which we connect with the designation of a system as living, and is thus a characteristic, necessary condition for living systems.
Thus, we consider ourselves entitled to apply these concepts in order, by means of them, to designate a certain relation existing in living systems between the external action, that is, the change of state of the surrounding environment, and the change of state of the system, whereby this relation has an exact physical significance.
There is no ground for setting apart the concept “irritability” in the form in which it is usually understood, as a property of living organisms in which the external action and the subsequent change of state show a certain disproportion, while the individual reaction chains are unknown, whereby, however, it is presupposed that this concerns only individual discharges, individual equalisations of potentials, which are determined only by the initial conditions. The more so since this property, as we showed above, is not only not characteristic of living systems, but does not at all correspond to those fundamental requirements which we made of the designation “living.”
Here it is fitting to point to one circumstance, very often encountered in the formation of concepts and the explanation of vital processes, which leads—as we shall yet often have occasion to show in further special examples—to the application of misleading concepts and to erroneous explanations.
This circumstance consists in this, that the course of the conspicuous phenomena which appear characteristic of vital systems is not carefully analysed by biologists, but is thoughtlessly compared with the course of purely physico-chemical phenomena, and therefore, in striving to subsume a phenomenon in living systems under the laws of physics and chemistry, they devise conditions which allegedly must exist in the organism in order that the course of phenomena in it should correspond to these laws already known from physics and chemistry.
For these characteristic phenomena, the necessary specific regularities are not sought out, and concerning the latter no suppositions are made that could be experimentally verified; rather, it is assumed in advance that the law of the phenomenon under consideration is in essence identical with the already known laws of physico-chemical phenomena. Suppositions are made not concerning the physico-chemical conditions that determine the peculiarity of the laws of vital phenomena. On the contrary, these laws are presupposed as known even before the study of vital phenomena, and suppositions are made only concerning the conditions which determine the course of these phenomena, if it is assumed in advance that the laws of these phenomena are already known. But since science explains the phenomena it studies only in so far as it establishes their laws, such a manner of “explaining” vital phenomena in essence explains nothing. As a result of this manner, the vital phenomena requiring explanation receive for the most part a very simplified explanation, not corresponding to the facts, and the biological concepts that were established for these characteristic phenomena are identified with the physico-chemical concept applied for their explanation. In consequence of this it is natural that these biological concepts seem superfluous and unfounded—not, however, because this is really so, but because they were in this manner deprived of their true sense.
In discussing various special vital phenomena and their theories, we shall often have the opportunity of adducing examples of this erroneous physico-chemical manner of explanation. Here it should only be pointed out that the concept of irritability, as we have seen, underwent the same fate. It was established as a biological concept for the unification of certain characteristic phenomena. Instead of subjecting it to a detailed analysis and investigating the physico-chemical conditions that must be fulfilled in these vital phenomena, they thoughtlessly compared them with the already known phenomena of discharge, whereby the biological concept of irritability received no physico-chemical sense, while losing, however, its biological sense.
The kind and direction of the changes of state of a living system in dependence upon the state and changes of the surrounding environment
Up to now we have discussed two properties characteristic of all systems which we designate as living, the presence of which is unconditionally necessary in order to designate a system as living. These properties, as we have seen, have a clear physical sense, and we therefore consider them necessary conditions which must be fulfilled and are fulfilled in every living being. We have, however, also seen that the presence of these two properties alone is not yet sufficient to designate systems as living. Evidently, to the designation “living” yet other requirements are made, to the discussion of which we shall now pass.
Our first requirement consisted in this, that living systems, with the surrounding environment unchanged, should not be in equilibrium, but should possess differences of potential which are equalised without external assistance, that is, they should be capable of performing work. The second condition required that, upon some action from without—consequently, upon some change of state of the surrounding environment—the system should perform work which would influence the changes of state called forth by this external action, and would change them. But both here and there we left open the question of in what work the capacity for work [работоспособность — rabotosposobnost′] should be expressed and in what the action of the system upon the externally induced changes of its state should consist. We have not yet established any requirements as to the kind, direction, and magnitude of that work which we required of living systems both with the surrounding environment unchanged and under changes of its state. We now intend, with respect to this point too, to make one requirement.
We assert that the material systems designated by us as living possess, in this respect too, a characteristic property, and we call living only such systems as possess this property. This property we shall formulate as follows: in order to designate a system as living, we require of it that it should expend its capacity for work, that is, its free energy [свободная энергия — svobodnaia energiia], which it possesses with the surrounding environment unchanged, always upon such work as leads, under the given surrounding environment, to changes in the conditions of the system that raise its capacity for work. In other words, there must exist in the system devices conducive to this, that the capacity for work of the system, even with the surrounding environment unchanged, should always be expended upon such works as raise the capacity for work of the system itself. In short, the capacity for work must be used in the interest of its own capacity for work.
Since a system is capable of work only when it is not in equilibrium, since at equilibrium there cannot occur in it, without external assistance, under unchanged external conditions, any changes whatever, we may formulate our requirement in yet this manner: in living systems there must always be devices which convert the capacity for work of the system, with the surrounding environment unchanged, into work against the equilibrium that would set in under the given surrounding environment and under the given state of the system.
By this we have set a condition with respect to the kind and direction of the work too, which must be used with the surrounding environment unchanged. In the case of external actions, that is, of changes of state of the surrounding environment, we find it obligatory, in our second requirement, that the system should perform work changing the effect of the external action. For the given case, with respect to the kind and direction of the work, we set the same requirement. Here we must recall that we set this requirement not unconditionally at every external action, but required only that such works, changing the effect of the external action, should in general be performed. Then our third requirement for this case will be expressed as follows: if an external action occurs upon a living system and the latter performs work influencing the effect expected from this action and from the initial state of the system, then the work of the system consists in such a change of the effect as a result of which, at the given external action, with the surrounding environment changed, the capacity for work of the system is raised; or, in other words, in the case where the system, at an external action, performs work, this work is directed against the equilibrium that would set in with the surrounding environment changed and the initial state of the system.
Briefly expressed, our third requirement consequently states: the work of living systems, under any surrounding environment, is directed against the equilibrium that would have to set in under the given surrounding environment and the given initial state of the system.
It should once more be noted that this requirement, like the second, in no way contradicts the laws of physics, in particular thermodynamics, for indeed by the state of the system—consequently, by the pressure, volume, temperature, energy store, entropy, and so forth, of its various phases, on the one hand, and the external conditions, on the other—the ensuing state of equilibrium is, plainly, determined unequivocally only when no devices within the system are presupposed that are connected in some manner with a change of state of the system or of its individual phases, or when concerning these devices, connections, concerning the properties of the boundary surfaces, and so forth, there exist exact indications.
But whether such devices exist, and in what relation they stand, is not to be discerned from the enumerated properties of the system. Our requirements, which we make of living systems in the sense of capacity for work and its application in a determinate direction, thus represent, properly speaking, conditions relating to the devices, relations, and so forth, which must be fulfilled in living systems.
The laws of thermodynamics in general only say that equilibrium sets in when with the given conditions of the system no change is compatible that would be accompanied by an increase or decrease of a certain magnitude of state, of entropy or of free energy. Whether equilibrium will set in at a determinate state or not—consequently, whether the system will be capable of work at a determinate state or not—depends on the conditions of the system. If, then, the processes occurring in the system and serving as its source of energy at the same time change the conditions of the system, if there exist in the system devices which conduce to the conversion of free energy into work that changes the conditions in the system itself, then it, in consequence of this work, under the same external conditions and actions and the same initial state, will remain capable of work, whereas without these devices and at the same magnitudes of state it would already have come into equilibrium. Thus, our third requirement means that in living systems work must always consist in a change of the structure of the parts of the system themselves.
We shall explain this by an example: let there be given a vessel (Fig. 1) with a movable piston impermeable to air; the vessel is partly filled with a solution of a combustible substance R at 20°; the piston is under a determinate pressure p, for example the pressure of the atmosphere. The substance R will enter into reaction with the oxygen of the air, whereby certain products of reaction R₁, R₂, and so forth, may be formed. Then the reaction will be obtained in the form of the equation: O₂ + R = R₁ + R₂ + … and, according to the law of mass action, equilibrium will set in if

where C_{O₂}, C_R, and so forth denote the molar concentration of the corresponding substances, and K a magnitude constant for the given temperature, the so-called equilibrium constant. If with this the products of reaction R₁, R₂ are not gaseous, then in the gas-space above the liquid O₂ will decrease, and consequently the pressure of the gas will be less; thus the piston will be advanced downward by atmospheric pressure until atmospheric pressure is also established within. With this the partial pressure of oxygen will decrease, and since the concentration of oxygen in the solution is determined by its partial pressure, then consequently C_{O₂} too will decrease until equilibrium is established in accordance with the cited equation. In the case where one of the products of reaction is, in burning, converted into a gas, for example CO₂, then it enters the gas-space, the partial pressure of O₂ will decrease still more rapidly, and equilibrium will set in still sooner. In this reaction a certain quantity of heat is liberated which, however, with this arrangement is not converted into any work, since the wall of the piston is permeable to heat, and the process proceeds isothermally.
Let us now imagine that the piston is connected with another similar piston situated in a vessel which is filled with gas or water vapour, and that this piston is loaded with a weight. The heat liberated in the reaction warms the gas or water vapour, whereby its pressure increases, the weight rises and will be transferred onto the first piston by means of, for example, a spring, which is held by means of a plate and is opened by the rising piston (K₁) (Fig. 2). By this the pressure on the first piston is increased, consequently the partial pressure of O₂ above the reaction mixture is raised, and consequently the concentration of O₂ too, and the reaction can proceed still further until equilibrium is reached by the law of mass action according to the equation cited above. This means that equilibrium will now set in not at the same state. Work has been performed by the system against the equilibrium that would have set in without this work; or, in other words, the system, at a certain quantity of oxygen in the air, is still capable of work, which it would not have been without this work under the same conditions.
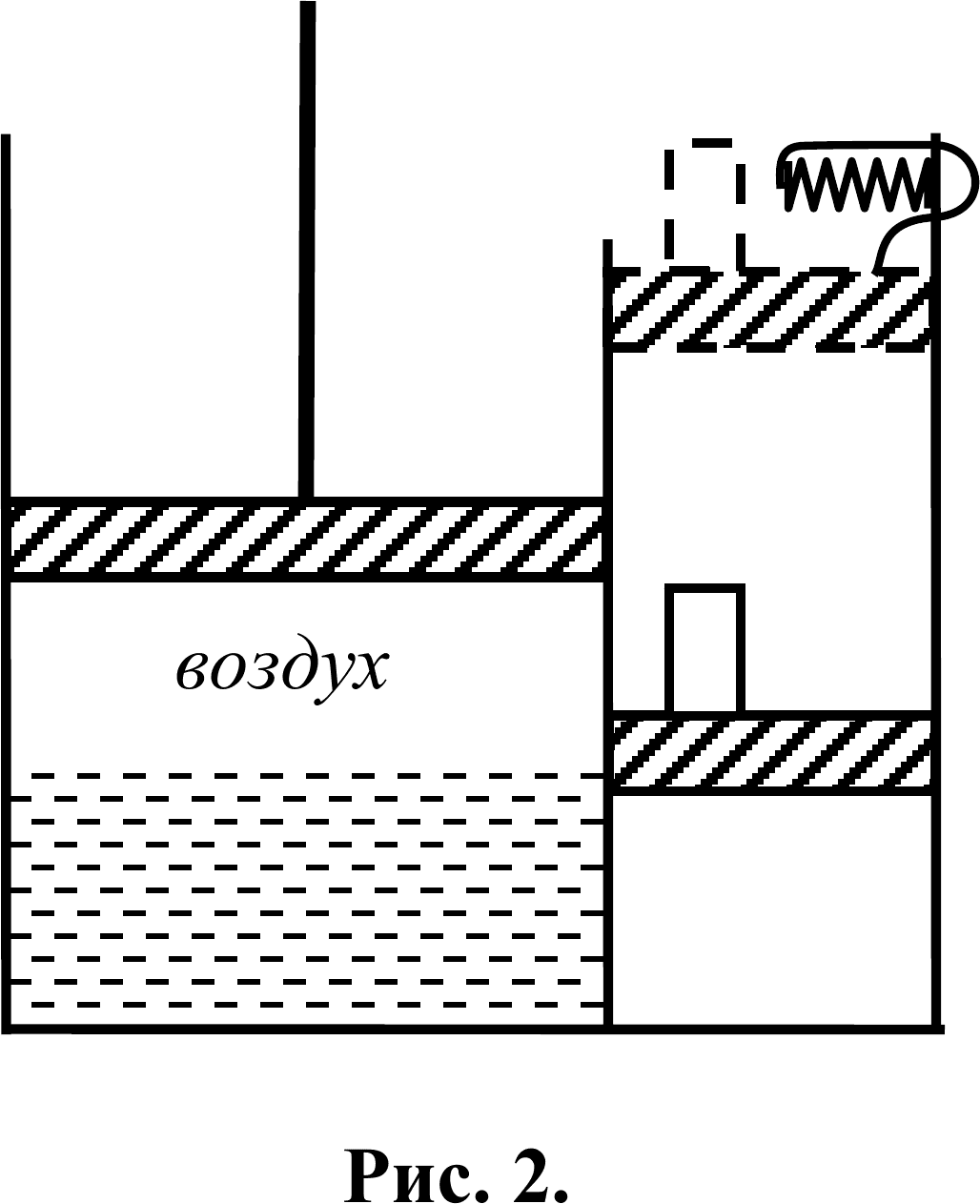
Whether the heat liberated in the reaction will be converted into work or not—this cannot be derived in advance from the above-mentioned magnitudes of state of the individual phases, such as: the partial pressures of the gases, the temperature, the concentration of the reaction mixture, the energy of the system, and so forth; it depends exclusively on whether the wall separating the two pistons is permeable to heat. Whether the work will be directed precisely against the equilibrium that would have set in without this work also does not depend on the given magnitudes of state of the system, but on the fact that the plate is opened by the rising cover; if the plate were placed above the spring, then, although work would be performed by the system and the weight would be raised, this work would not change the ensuing equilibrium.
From this example it is clear that the state in which a system, at a certain surrounding environment, comes into equilibrium, and the work performed against the equilibrium, depend in fact not only on the thermodynamic magnitudes of state of the individual phases, but also on the conditions of the system.
We have deliberately chosen such a graphic example, based on primitive mechanisms, in order, by comparison with the processes of respiration, to make clear also to biologists and physiologists as graphically as possible the significance of all these considerations for biology. It should be mentioned that the question of deviations from the law of mass action, or of the establishment of a so-called “false equilibrium” not answering to this law, has again become highly topical in physical chemistry. These deviations from the equilibrium expected on the basis of the law of mass action and the laws of thermodynamics set in within a homogeneous system; for example, in a mixture of P₄ with water vapour. Here the vapours of phosphorus and oxygen react with one another at a great rate. But as soon as, in consequence of this reaction, the concentration of oxygen falls to a determinate magnitude, the rate of reaction suddenly falls to zero. The reaction halts long before the onset of equilibrium.[Author’s note:] N. N. Semenov, Chain Reactions, p. 37, Goskhimtekhizdat, Leningrad, 1934.
In these cases we are dealing not with macroscopically coarse parts of the system acting as transformers, as in our previous examples, but the principle here too remains the same. These cases are explained by means of so-called chain reactions, in which the free energy of activated molecules activates other molecules even before their entry into reaction or before their return to the initial state. Thus, here too we are dealing in principle with the same phenomenon, namely, here the free energy of the system changes the structure of the parts of the system themselves, and neither directly performs work against the surrounding environment nor is directly liberated in the form of heat. It is therefore understandable that Semenov, precisely in connection with these cases, points to the insufficiency of thermodynamics for the determination of the position of equilibrium. He writes: “A system that is in true equilibrium cannot, under the action of internal forces, react and thus disturb the equilibrium. Thermodynamics forbids macroscopic disturbances of an established equilibrium. Further: thermodynamics requires that all occurring processes should proceed in the direction of approach to equilibrium. It would, however, be incorrect to assert that thermodynamics requires that in a system not in equilibrium there should necessarily always occur processes of approach to equilibrium.” “The above considerations may be formulated thus: all changes forbidden by thermodynamics do not in actuality occur, but if thermodynamics permits some change, it may also not occur.”[Author’s note:] Semenov, ibid., p. 33. As we have seen, what is characteristic of living systems is precisely that they, at the expense of their free energy, perform work against the expected equilibrium, and thus we are dealing not with a contradiction of the laws of thermodynamics, but with other laws, consisting, among other things, in this, that what is permitted by thermodynamics regularly does not set in. Later, in the chapter on the general theory of living matter, we shall see that in living systems too we are, in the final account, dealing not with such coarse, macroscopic mechanisms and structures as those cited in the previous examples, but with non-equilibrium structures of the molecules themselves and with work that maintains this structure of the molecules. Where chain reactions are present, where consequently the free energy of the system partly performs work that changes the structure of the parts of the system themselves, that activates molecules, there too the regularities and concepts of physics and chemistry, in particular of thermodynamics, are insufficient for the investigation of the regularities of the chemical dynamics of these processes. “The question here comes down to the discovery of new dynamic characteristics of the initial substances, that is, of those properties which a substance manifests during one or another of its transformations.” “We must only remember that the construction of a theory of chemical dynamics is bound up with a number of profound reconstructions in the very system of our sciences, which are for the time being adapted in the main to the solution of static or, at best, equilibrium systems.”[Author’s note:] Semenov, ibid., pp. 12–13.
If, then, already in chemical dynamics, in connection with the formation of chain reactions, thermodynamics, adapted to equilibrium states, is insufficient, then it is clear that where work against equilibrium, and consequently against the parts of the system themselves, appears as the fundamental property and the general regularity, as is the case in living systems, there we stand before other, new regularities, which can no longer be represented by means of corrections introduced into the old concepts, since here it is precisely these deviations that become the regularities.
In our last requirement we said that the work of living systems is directed, under any surrounding environment, against the equilibrium that would have to set in under the given surrounding environment and under the given initial state of the system. Thus, in our last example we adduced a system that converted its free energy into work which raised the capacity for work of the system.
Does this system, then, correspond to our requirement which we set for living systems? No, far from it! In the first place, the system is so constructed that it can convert the liberated heat into work only at the given atmospheric pressure, but cannot perform similar work with the surrounding environment changed, for example at a lowered atmospheric pressure: the piston K will then be unable to descend, since the reaction, in accordance with the law of mass action, will cease sooner, the heat produced will be insufficient to raise the piston K up to the plate, and the whole arrangement will be useless; no work against the expected equilibrium under this surrounding environment will be performed. This would be possible only in the case where, at the expense of the heat of reaction, work were performed that would change the conditions of the system. The system, so to speak, can no longer adapt itself to the diminished external pressure. Or—according to our above-cited definition of stimulation as a change of state of the surrounding environment which calls forth excitation, that is, an effect proceeding not as one would expect on the basis of the change of the surrounding environment and the initial state of the system—for the given system the decrease of external pressure represents no stimulation whatever.
Our third requirement states that work against equilibrium is possible under any surrounding environment. But this is only one side of the matter. We required work not only against the equilibrium that would set in under any surrounding environment, but also against that which is determined by this environment and the initial state of the system. In the construction we adduced as an example there are given not only the magnitudes of state of the individual phases of the system and the magnitudes of state of the surrounding environment, but also the initial conditions of the system with all the relations and conditions of the bordering surfaces (boundary conditions). With such an initial state and with the surrounding environment unchanged, there must set in precisely that equilibrium which sets in after the transfer of the weight onto the piston K. In this case our requirement would be fulfilled only in the case where the heat were converted into work directed against this latter equilibrium. This again is possible only when work is applied in the course of the process itself for the change of the conditions of the system.
We see that a necessary condition for the fulfilment of our third requirement is that the work of the system should be applied to the change of the conditions of the system, consequently to the change of structure, to the creation of a difference of potentials in the system, and not only to the performance of external work. The raising of the weight by means of the piston in our example also creates differences of potential, but not at the expense of a change of the structure of the system, rather at the expense of the performance of work against the surrounding environment, that is, in the form of external work. The two pistons are here connected in a purely external manner. Let us now see what would have to occur in our last example in order that the construction should correspond to those requirements which we make of a living system.
In the first place, with the surrounding environment unchanged, the conditions of the system would have to change in such a way that there should set in not that equilibrium which we expected under the given initial conditions described by us above; that is, in order that in our example, after the descent of the weight onto the piston K, equilibrium should not set in, one must employ the heat of the chemical reaction liberated in this for work which would change the conditions of the system in such a way that the system should still remain capable of work.
This can, of course, be pictured in the most various ways, namely, at the expense of this heat of reaction the conditions of the system must change, which will either raise the partial pressure of oxygen or else diminish the concentration of the products of reaction. The first can be attained only in the case where, at the expense of the heat of oxidation, a further compression of the gas occurs through a change of the conditions of the system, for example a change of the elasticity of the walls in the case where they are elastic. The second could occur only on condition that the heat of oxidation, in consequence of a change of the conditions of the system, calls forth a diffusion or a current of the products of reaction, for which work would also be required, namely the creation of differences of potential, of a difference of pressure of concentration or of hydrostatic pressure in the system.
If we now consider the living organism of a man or of an animal breathing with lungs, which, as is known, has as its principal source of energy likewise the energy of oxidation of the oxygen of the air, and where these processes of oxidation proceed chiefly likewise in a liquid medium, in colloidal aqueous solutions, then we shall see that the requirements set forth above are fulfilled here precisely in the manner described. To the air-space above the piston there corresponds here the air-space of the lung, to the wall of the cylinder—the thoracic cage, to the liquid reaction mixture—the tissues of the body. And we know that a part of the energy of oxidation is converted into work changing the elasticity or tension of the walls of the cylinder, that is, of the muscles of the thoracic cage—this is the work of respiration. Another part of the energy of oxidation is used in order to produce a current of the products of reaction—this is the work of the heart; a part of the energy is applied to the removal of the products of reaction by means of the creation of differences of concentration in the system—this is the work of the kidneys. All these devices serve, consequently, in the animal organism breathing with lungs, in order to fulfil the requirements set forth by us: to perform work at the expense of the free energy of the system which would so change the conditions of the system that it should remain capable of work, consequently, that under the existing external conditions equilibrium should not set in. In lower animal organisms—in the unicellular ones—and in plants, the same requirement is fulfilled by means of quite other mechanisms and along quite another path. But these organisms too possess devices in order to be able to perform this work against the expected equilibrium upon a change of the surrounding environment. We know, taking the former example, that animals breathe more intensely at a diminished pressure of oxygen, that is, the work for the creation of differences of elasticity of the thoracic cage becomes more intense; the same holds also for the creation of differences of hydrostatic pressure, that is, the work of the heart, and so on, so that the intensity of oxidation is to a considerable degree independent of the pressure of oxygen in the surrounding environment. That unicellular organisms too possess such devices, which give them the possibility of performing work against equilibrium upon a change of the surrounding environment, follows from the fact that in them too there exists a similar broad independence of the intensity of oxidation from the pressure of oxygen. We see, however, also that all these devices consist not in a direct conversion into external work, but have as a necessary presupposition work directed to the change of the structure of the parts composing the system, in the sense of the preservation of the capacity for work of this structure.
From the above-cited explanations and examples it follows that, whereas the first requirement corresponds approximately to the requirement of the properties of a wound-up machine, the second requirement—to irritability and excitability, the third requirement corresponds to those properties of living beings which are usually designated as adaptability, purposiveness, regulation, wholeness, and are regarded as characteristic of living beings.
On the other hand, we have seen that the second requirement presupposes the fulfilment of the first, and the third—the fulfilment of the second, because only a system not in equilibrium—consequently, a system whose free energy can diminish without a change of external conditions—can perform work changing that effect of the external action which one would expect as a result of the initial conditions, and thus be excitable. And only an excitable system in such a sense can always perform work against equilibrium, consequently fulfil the third requirement. This, however, means that the third requirement, if all its consequences are taken into account and its significance is fully made clear, contains within it all three requirements.
Now we must again pose the question: shall we in fact designate a material system that corresponds to our third requirement as a living system, or are there such systems as satisfy the third requirement but which we nonetheless do not designate as living?
It is plain that theoretically the existence of such systems is quite possible, which correspond to this requirement and which we shall nonetheless not designate as living. On the other hand, we cannot verify whether all existing living beings correspond to our third requirement. We may, however, lay at the foundation of our biological investigations the assumption that all living beings answer to this third requirement and that the theoretical possibility of the existence of a non-living system which nonetheless corresponds to our requirement is in principle excluded. We shall then obtain a universal law of biology, which states: “all and only living systems are never in equilibrium and, at the expense of their free energy, continually perform work against the equilibrium required by the laws of physics and chemistry under the existing external conditions.”
Having accepted this law as valid, we can then apply it in every individual case, that is, at every vital phenomenon of every living being, whereby, proceeding from its correctness, we can indicate that the vital phenomenon under consideration in fact represents a special case of this law and does not contradict it. Such a general law, in so far as it is correct and leads in every individual case to true conclusions not contradicting the facts, and is therefore always set at the head of every investigation, is called a principle. Thus, for example, one speaks of “d’Alembert’s principle,” “Hamilton’s principle,” “the principle of inertia,” and so forth, in mechanics, of “the principle of the conservation of energy” in thermodynamics, and so on.
We shall designate this principle as the “principle of stable non-equilibrium” [принцип устойчивого неравновесия — printsip ustoichivogo neravnovesiia] of living systems. This designation clearly expresses the sense of the principle and the marks of living systems characteristic from the standpoint of thermodynamics. Just as stable equilibrium is characterised by the fact that, having been disturbed, it always sets in anew, so too in living systems the non-equilibrium state is constantly preserved and possesses all the marks of stability. Our principle also expresses in brief form a characteristic property of living systems, since we know not a single non-living system in which the non-equilibrium state would possess the marks of stability.
The principle we have formulated speaks only of the general behaviour of living beings and of the direction of the processes proceeding in them; it, however, contains no quantitative characteristics. Therefore we must supplement it still further, so that it should receive a quantitative expression. We assert that a living system always converts its entire free energy into work against the expected equilibrium. This expression is quantitative, and its correctness may be experimentally verified by means of measurements.
The quantitative principle must, of course, be expressed by a formula. Before doing this, we should like to clarify somewhat the sense of this quantitative expression. First of all, from this expression it follows that a living system in general performs no other work whatever besides the work against equilibrium. Indeed, if it applies its entire free energy to this work, then it can no longer perform any other work. If we designate the work which is directed against the expected equilibrium, or, as we have seen, leads to the reproduction of differences of potential in the system, consequently to the preservation of its capacity for work, as regulating activity, then from the quantitative principle it follows that there exist only regulating vital activities, or that every vital activity is regulating. Then from the quantitative principle it follows that if we isolate a living system under certain constant conditions—consequently, do not permit any external action, that is, any influx of energy—then, although the living system will also come into equilibrium, the equalisation of all the differences of potential will proceed not as in a wound-up machine with unchanged conditions of the system, but otherwise—namely, the entire free energy of the system will be directed to the retardation of the equalisation. This second consequence of our quantitative expression can be expressed by a formula and is accessible to direct experimental testing.
We shall now make an attempt to give this formulation in an elementary manner. Let us imagine that we have an isothermally closed system, that is, one whose wall is permeable to heat, whereby the process proceeds so slowly that the temperature can be regarded approximately as constant. The second law of thermodynamics states that in this case, first of all, equilibrium will necessarily set in; this means that such a system cannot infinitely perform work, since this would contradict the second law, according to which it is impossible to build a system performing constant external work at the expense of heat. The maximal work that can be extracted under such circumstances from the system is the measure of the free energy of the system, and from the second law it follows that equilibrium will set in at such a state in which the free energy, under the given conditions of the system, can no longer diminish, consequently, in that state in which the free energy is a minimum. Our principle thus shows that in an isothermally closed living system the total free energy of the system is transformed into work which produces such changes of the conditions of the system that this minimum takes on the smallest possible magnitude, not only relatively but also absolutely.
From this it follows that the free energy in an isothermally closed living system at the onset of equilibrium will be less than in a non-living system, although in both, at the outset, the sum of the differences of potentials, expressed in absolute magnitudes, would be one and the same. Under these conditions the difference in free energy between the living and the non-living system at the onset of equilibrium is exactly equal to the magnitude by which the work of the living system in the course of the whole process of equalisation exceeded the work of the non-living system.
Thus, if we designate the free energy of the living system at the onset of equilibrium by F, the sum of the differences of potential, otherwise the factors of work—such as difference of pressure, difference of concentration, electrical difference of potential, and so forth—we designate by X, the changes called forth by these factors—by ∆x, and the time during which this change occurred, by ∆t, and these same magnitudes, but for the non-living system, we designate by the same letters but with the addition of the sign ′—then we obtain the following formula:
With this, the summation on the right-hand side is to be understood thus, that we divide the equalising process into intervals of time ∆t and successively, for each interval of time ∆t, multiply the corresponding factor of work X and the rate of the change called forth by the factor of work by one another and by the time ∆t, and successively sum up these products. Thus we obtain, both for the non-living system and for the living system, the total work performed by them during the process of equalisation; the difference then gives the work of the living system against the equalisation. This difference must be equal to the difference of free energies, which is precisely what our quantitative principle shows, namely, that the total free energy is converted into work against equilibrium. Since the factors of work, plainly, change at each moment, then, taking finite intervals of time ∆t, we obtain for the expression of the work only an approximate magnitude. The intervals of time should be taken smaller and smaller, and the limiting number of the sum which we obtain if we continue to diminish the intervals of time will be the actual magnitude of the work. This limiting number is designated in mathematics by the so-called integral sign, and the exact formulation of the principle in the language of higher mathematics will then be as follows:
This is the mathematical formulation of our quantitative principle of biology.
If we look more closely into the formula, then we shall see that the difference of the two integral magnitudes, otherwise of the magnitudes of the sums, rests upon the fact that the conditions of the system, or, expressing it in the language of mechanics, the constraint or systemic forces themselves, represent in living systems other functions of time than those in non-living systems, and precisely in consequence of the fact that they are forces obstructing the equalisation. The constraint forces, representing functions of time, perform work which is directed against the process of equalisation, whereby the source of this work lies in the living system itself—therefore towards the end it must possess a smaller free energy, and precisely by so much smaller as the greater work the forces of the system performed in the equalisation. In the final account the matter in fact comes down, as was to be expected, to the work needed for the preservation of the conditions of the structure.
Two small examples will explain this to us: the difference in the integral expressions, which thus expresses the work of the forces of the system, consequently of the conditions of the system, that is, that work which is needed in order to change the conditions of the structure in accordance with time, requires that in living systems x should be another function of . If we take, for example, the process of diffusion, then we must take for x the difference of concentration c₁ – c₂, that is, the difference of concentration on the two sides of the membrane, and for the rate at which the substance under consideration passes through a unit of surface of the membrane, consequently the quantity passing in a unit of time; we know, however, that for this case the following equation is valid:
consequently:
where D denotes the diffusion constant, that is, a material constant, a structural property of the wall, which enters into this formula as a constant independent of time. In living systems, however, the ratio of the rate of diffusion to the existing difference of concentration (c₁ – c₂) is not constant, that is, the material constant is in dependence also on time itself, and precisely in such a manner that it changes owing to the work of retarding the equalisation. The work that is needed for the change of these properties of the structure is given precisely by the difference of the two integrals. In the equalisation of electrical differences of potential, the ratio to x likewise characterises the material constant, consequently a property of the structure. Since here x is equal to the electromotive force v and dx is equal to de, if e denotes charge and , consequently , then, however, , that is, the intensity of the current, and if R denotes resistance, then we obtain:
where R denotes resistance; that is, the ratio characterises the conductivity of the material.
In general it may be said that whereas the processes of equalisation in non-living systems are determined by the immediate properties of the structure, in living systems the matter stands otherwise, because the properties of the structure themselves change during the process of equalisation; on the other hand, as we shall see in further examples, the driving forces of the very processes of equalisation, that is, the factors of work (x), show a different dependence upon the rate of equalisation , inasmuch as the former can during the equalisation be newly formed or diminished; on the other hand, the rate of equalisation is connected with the factors of work otherwise than is the case in non-living systems, which plainly depends on certain peculiarities of the structure. The chief difference consists, first, in this, that this different dependence is characteristic of living systems, inasmuch as it is always directed against equalisation, and, secondly, in this, that for the work which is necessary for the change of this dependence, the entire free energy is in fact expended.
On the relations to dynamic equilibrium, to Le Chatelier’s principle, and on the significance of the structure of living systems
In the preceding chapter we have analysed in detail the content of our qualitative and quantitative principle, which we set at the foundation of our entire exposition. This detailed explanation was unconditionally necessary in order to understand the exact physical sense of the principle and to become familiar with it. Further proofs of this principle and its application will outline still more clearly its biological, as well as its physical, sense. Before passing to the proofs and applications, it is important to make clear to ourselves yet in what relation the system characterised by us stands to so-called “dynamic equilibrium” [динамическое равновесие — dinamicheskoe ravnovesie], and what relations exist between our principle and so-called Le Chatelier’s principle [принцип Ле Шателье — printsip Le Shatel′e], since on these two points great misunderstandings often occur. In close connection with this stands the question of the significance of the structure of living systems.
Dynamic equilibrium
Very often it is asserted that a living system is in dynamic equilibrium. What is meant by this? First we must make clear to ourselves what is understood by dynamic equilibrium in physics or in physical chemistry.
Let us see what Nernst says about this:[Author’s note:] Nernst, Theoretische Chemie, Stuttgart, 11th–15th edn., p. 523, 1926.
“The view that in equilibrium there exists no absolute inactivity between the reacting substances, but that rather, strictly speaking, the reacting component parts are unceasingly subject to further mutual action, and that here precisely only the mutual exchange is balanced, has fundamental significance for the elucidation of changes of substance in general. It is usually said that equilibrium in this and in analogous cases is ‘not static, but dynamic.’ “Thus, the equilibrium between water and water vapour, according to Clausius, is to be understood approximately not as though here there occurs neither evaporation of liquid water nor condensation of the gaseous, but rather both processes continuously take place at equilibrium too, that is, at the contact of saturated water vapour with water; only at any moment there passes through a unit of surface of the liquid water an equal quantity of water molecules in both the one and the opposite direction.”
Consequently, by dynamic equilibrium is in general designated such a state of a system as remains unchanged in the absence of external action; this system is therefore incapable of work, that is, its free energy under the existing conditions cannot diminish, but such a state of equilibrium is obtained in consequence of the fact that within the system every change is annulled by a simultaneous change equal in magnitude in the opposite direction. Thus, for example, the equilibrium that sets in at a reversible chemical reaction in a test-tube represents such a dynamic equilibrium; when, for example, one mole of alcohol and one mole of acetic acid mutually act upon one another, then the reaction proceeds according to the equation:
and there are thus obtained acetic-ethyl ester and water. This reaction proceeds until chemical equilibrium sets in, that is, until we obtain a final unchanging combination, which will be the following: 1/3 mole of alcohol, + 1/3 mole of acetic acid, + 2/3 mole of ester, + 2/3 mole of water. This unchanging mixture, that is, a constant concentration of each individual one of the 4 component parts, is obtained in consequence of the fact that at each moment there is formed the same quantity of molecules of alcohol and molecules of acetic acid as of molecules of water and ester; therefore this equilibrium is also called “statistical equilibrium.” In the thermodynamic respect, consequently in respect of capacity for work, this equilibrium is completely identical with static equilibrium. A salt solution of a determinate unchanging concentration represents in essence precisely such a dynamic or statistical equilibrium. If we take a glass of water and throw into it a certain quantity of salt, then at the first moment no equilibrium will be obtained, since the concentration of the salt will not be the same in every part of the water, and until then changes of concentration by means of the diffusion of the salt molecules will occur spontaneously, without external action or ions, until the concentration of the salt is everywhere the same. Then no change of state will occur in the system any longer—the system will be in equilibrium and will be incapable of work, whereas before this, until uniform distribution had set in, it was not yet in equilibrium and was capable of work, since the diffusion of the molecules from places of higher concentration to places of lower concentration can be converted into work by means of suitable mechanisms and devices. The final equilibrium of uniform concentration in essence represents also a “dynamic” or “statistical” equilibrium, since the molecules or ions of the salt are not in repose, but in constant, non-uniform Brownian motion, and if we mentally cut out a part of the space of the water, then molecules will constantly issue from it and enter into it, but in every not too small interval of time there will always issue the same quantity as enters. Nonetheless, this system will not be capable of work, and the Brownian motions, which represent thermal motions, can by no means be converted into work without external assistance. Thus it is clear that living systems are never in that state which in physics is sometimes designated “dynamic” equilibrium. The point is that the living system, as we have made clear above, remains capable of work even without external action, that is, when we isolate it from external actions, and in this case there occur in it changes in which the free energy diminishes—thus energy is liberated which can be converted into work. One can adduce another example of so-called dynamic or statistical equilibrium, when the system is not closed and when it is capable of work. Such a case is represented, for example, by a waterfall. The comparison of a living organism with a waterfall is very often applied. How incorrect it is we shall now show.
A waterfall preserves approximately its form, can constantly perform work—for example, drive a mill—at the expense of the difference of potential which exists between the upper and lower level of the water and which is constantly equalised and converted into kinetic energy, whereby ever-new quantities of water fall from above downward. This state can be called dynamic, or statistical, equilibrium, since at each moment of time the same quantity of water flows in as flows out.
First of all, it is clear that the existence of this dynamic equilibrium is bound up with the existence of a condition independent of the system—consequently, of the waterfall—an external condition, namely with the existence of a source supplying water. And the capacity for work of the system is also bound up with this external condition. If we isolate the system under unchanged external conditions, which comes about with the shutting off of the source supplying water, then we obtain in the waterfall a system which is not in equilibrium, whether dynamic or static, similar to a wound-up machine which very quickly stops: the water diminishes, and equilibrium sets in. At the expense of the kinetic energy of the water that appears in this, however, no work against equilibrium is performed; for the preservation of the capacity for work of the system—consequently, of the waterfall—no difference of potential is created, no source is created, consequently the overlying reservoir of water is not filled.
Thus the “dynamic equilibrium” of the waterfall exists only so long as, independently of the performance of work by the waterfall itself, there exists a source of energy—the potential energy of the overlying reservoir of water or of the spring. On the contrary, the state of capacity for work of living systems is obtained, according to the principle adduced by us above, at the expense of the work of the system itself. The capacity for work of living systems is obtained not directly thanks to the influx of energy from a source of energy existing independently of the system. The living system creates the source of energy, the differences of potential, at the expense of the free energy existing in the system. This means that it works against the equilibrium of the system under the existing surrounding environment. The point is that the source of energy for animal organisms is, as we know, the chemical energy of nutrition, which is liberated by the splitting of food.
Even if we do not take into account the work of obtaining and seizing food, which is performed at the expense of the free energy of organisms, we must reckon with the work of digestion, the entry of dissolved food, usually accompanied by osmotic work, and also the process of blood circulation, that is, that colossal work of the heart, without which the supply of food to the cells is impossible. The oxygen necessary for the organism does not, by the law of diffusion, enter the living organism without the work of the latter, but is sucked in, as, for example, in man, with the participation of the system itself, for which is needed that great work which the respiratory muscles perform. These relations hold not only in higher animals and multicellular organisms, but in every living system, down to the most primitive unicellular ones inclusive. They all have corresponding mechanisms which, at the expense of the free energy of the system, use the sources of energy of the surrounding environment for the preservation of the capacity for work, that is, against the onset of equilibrium under the existing external conditions. In the “dynamic,” or statistical, equilibrium of the waterfall and of analogous non-living systems (flame, and so on) the source of energy for the preservation of the capacity for work, namely of the non-equilibrium of the system, lies outside the system; in living systems, on the contrary, within it. As, moreover, follows from the considerations adduced above, the waterfall would differ in principle from living systems even in the case where there were devices permitting its kinetic energy to be transformed into physical work, thanks to which a certain quantity of water would again be raised upward: for this would not be the work of constraint forces. In order that this fundamental difference should vanish, the difference of levels, thanks to which alone the waterfall is possible, would have to be created or maintained by the waterfall itself. This is sufficient to show that, in accordance with our demarcation of living systems from non-living ones and our principle, living systems have no relation whatever to so-called “dynamic equilibrium.” The conception that a living system is in dynamic equilibrium is physically at bottom incorrect and therefore leads biologically to erroneous conclusions not corresponding to the facts.
Le Chatelier’s principle
Here it should be mentioned further that our principle, according to which living systems, upon changes of state of the surrounding environment, always perform such work as is directed against the equilibrium expected under the given surrounding environment, has nothing in common with so-called Le Chatelier’s principle. Le Chatelier’s principle states:
“Every change of one of the factors of equilibrium modifies the system in that direction in which the factor under consideration undergoes a change opposite to the initial one.”[Author’s note:] See Nernst, l.c., p. 759.
This principle includes van ’t Hoff’s principle of mobile equilibrium (Principe de l’équilibre mobile), according to which a rise of temperature of a chemical system at constant volume displaces the equilibrium in that direction in which the reaction proceeds with absorption of heat. It also contains the following rule: a rise of pressure at constant temperature shifts the equilibrium in that direction in which the reaction is bound up with a decrease of volume.
For the first principle there serves as an example the fact that the tension of vapour rises upon heating, since evaporation is accompanied by absorption of heat. As an example for the second proposition we shall point to the fact that the compression of ice conditions its conversion into water, in consequence of the fact that the specific volume of water is less than the volume of ice; thus, the conversion into water is accompanied by compression.
Le Chatelier’s principle contains within it also a general indication of in what direction a system will change upon a change of the surrounding environment; it says that the change in the system will occur in the direction opposite to the change of the surrounding environment; consequently, an influx of heat leads to a reaction lowering the heat, a rise of pressure—to a reaction lowering the pressure (compression), and so forth.
Our principle shows that a living system, upon a change of the surrounding environment, will perform such work as is directed against the equilibrium expected under the given changed environment and under the unchanged conditions of the system.
Between the two principles—Le Chatelier’s principle and the fundamental biological principle established by us—there exists, thus, an external resemblance, consisting in this, that both of them contain a general indication of in what direction the reaction will occur, that is, the change of state of the system upon some change of state of the surrounding environment. Moreover, both principles say that the change of state of the system is directed in a certain sense against the change of state of the surrounding environment. The physical sense in both cases, however, is quite different and has no relation to one another. Notwithstanding this, it is sometimes considered that between these two principles there is something in common, or that the behaviour of living systems upon changes of state of the surrounding environment should be derived directly from Le Chatelier’s principle.
This erroneous analogy, just like the analogy of dynamic equilibrium, entails undesirable consequences, since it is physically incorrect and leads to erroneous conclusions not corresponding to the facts. To understand this, one must bear in mind the following. Le Chatelier’s principle relates to systems that are in equilibrium, and the change of state, that is, the reaction of the system, which the principle requires upon a change of the surrounding environment, leads precisely to the equilibrium expected under the given surrounding environment; in other words, the principle indicates at precisely what direction of reaction equilibrium will set in under the given new surrounding environment. Our principle relates to systems not in equilibrium, and the change of state, otherwise the reaction of the system, which our principle requires upon a change of the surrounding environment, consists in work against the equilibrium expected under the given surrounding environment, consequently precisely against that change which one would expect by Le Chatelier’s principle if the systems were in equilibrium.
Le Chatelier’s principle is properly a consequence of the second law of thermodynamics and can be derived from it. It can be applied to living systems too, like the second law, and does not contradict our principle of living systems. Our principle proceeds from the validity of the second law and of Le Chatelier’s principle, and it is precisely by means of them that it is possible to say in advance what changes of state would have to occur and at what state equilibrium would have to set in, consequently what work against it must be performed by a living system and at the expense of what devices and mechanisms existing in it. To the second law and to Le Chatelier’s principle our principle has no direct relation: it is not identical with them, but neither does it stand in contradiction with them; it already presupposes their validity and in essence gives certain indications concerning the conditions of structure and the devices of living systems.
Chapter 2: The Free Structural Energy of Living Systems and the Principle of the Work of Systemic Forces
On the basis of the internal conditions, characteristic of all living systems, under which changes of their state occur, and on the basis of the form and direction of these changes, we arrived in the preceding section at a general law of motion of living matter, which we named the principle of stable non-equilibrium. In so doing we saw that the validity of this law depends on the presence of determinate conditions of the system and of their changes, and that thus our law already, to a certain degree, presupposes or contains within it certain general assertions concerning the structural peculiarities of living matter and their changes. To this same general law of motion one can also arrive from the standpoint of these structural peculiarities of living matter, basing oneself on certain other empirical data concerning the change of states of living systems. Let us attempt to find these general laws of structural peculiarities, which serve as presuppositions for our principle of stable non-equilibrium. We shall set out for this purpose from the results of the classical experiments of Rubner, in which he proved the validity of the first law of thermodynamics for living beings too. As is known, Rubner showed by means of direct calorimetry that the quantity of heat emitted by an animal is equal to the heat of combustion of the food taken in by the organism, outside this organism (in a calorimetric vessel). This result signifies, however, considerably more than simply a confirmation of the first law of thermodynamics for living beings; it confirms this law in a quite peculiar form.
To what extent, then, and in what form is the first law of thermodynamics confirmed here? This law states, as is known, that upon the transition of some system from one state into another the sum of the work performed and the heat depends only on the initial and final state and does not depend on the path traversed in this transition. Mathematically this is expressed by the formula:
where and denote the entire energy of the system in the initial resp. in the final state, the mechanical work performed, and the quantity of heat given off. How, then, is this law to be applied to Rubner’s experiments? We have nutritive substances and subject them to certain chemical changes, burning them in a calorimetric vessel, whereupon they pass into a new state, namely into the final products of combustion: H₂O + CO₂. In this transformation in the calorimetric vessel our system, that is, the nutritive substances, performs no work, consequently is equal to zero, and therefore:
when a quantity of heat equal to is given off. In other words, the quantity of heat given off is a measure of the difference of the quantities of energy in the initial and final state, that is, before and after combustion. Rubner’s experiments on animals show that if the same nutritive substances are burnt to the same final products not in a calorimeter, but in the organism, then the same quantity of heat is given off, provided the organism performs no work against the environment. This of course confirms the law of independence from the path traversed in changes of state of a system. But of what system? Of the system of nutritive substances! And this only on condition that in their transformation in the organism is likewise equal to zero, that is, that at the expense of the energy of the food no work is performed in the organism either. But this is already something quite other than a confirmation of the first law of thermodynamics for organisms or living systems. Rubner’s experiments confirm the first law of thermodynamics not for living systems, but for nutritive substances in their transformations within the living organism, and this only on condition that in the living organism no work is performed at the expense of the energy of the means of nutrition. Conversely, if we set out from the validity of the law of the conservation of energy for living organisms too, then on the basis of Rubner’s experiments we must arrive at the conclusion that the organism performs no work at the expense of the energy of the nutritive substances. The coincidence between the quantity of heat in the calorimeter, on the one hand, and in the organism, on the other, is of course not accidental and proves the validity of the first law of thermodynamics. But we know, after all, that the principal and even almost exclusive source of energy for the organism is the chemical energy of the food which it takes in from the surrounding world. Therefore we are compelled to conclude that the organism performs no work at all, which evidently contradicts the facts.
Even leaving out of account the work performed by the organism with respect to the environment surrounding it, and considering only the organism in complete repose, we observe in the animals with which Rubner worked a great deal of work performed within the organism. It is enough to recall the work of the heart for the maintenance of the circulation of the blood, the work of the respiratory muscles, necessary for respiration, the work performed by the kidneys in concentrating the substances subject to excretion, and so forth. From Rubner’s experiment it follows that the differences of potential evoked by work within the organism are not preserved, but are again equalised, that is, that the work performed at the expense of chemical energy is never preserved within the living organism in the form of potential energy. Otherwise it could, of course, not immediately appear anew in the form of heat. But whether the difference of potential created in the system is preserved, that is, whether the potential energy of the system diminishes or not—this depends exclusively on the systemic conditions of the given system, that is, on its build, the structure of the system. If, for example, the potential energy of a spring is increased thanks to work from without (stretching), then the preservation of this energy in the form of potential energy of tension, or its immediate equalisation, will depend only on whether the end of the spring, in virtue of determinate structural conditions (for example, attachment to a fixed hook), remains in its new position or not. In the first case the stretched spring will represent a state with greater potential energy, that is, with greater capacity for work, but one that is in equilibrium; in the second case, however, we shall have the same state with the same potential energy, for the obtaining of which the same quantity of work must be expended, but it will not be in equilibrium, that is, it cannot be preserved under the given conditions of the system. Let us, for clarity, adduce one more example. A vessel A is separated from a vessel B by a semipermeable partition, vessel A being provided with a tube. In both vessels there is an aqueous solution of some substance. If now, by means of some work, a difference of concentrations is obtained in the two vessels—the concentration being, for example, higher in vessel A—then, as is known, the water in the tube will begin to rise, until the hydrostatic pressure of the column of water becomes equal to the osmotic pressure, that is, until equilibrium sets in. In this case the work necessary for the raising of the water is performed directly by the osmotic forces, while the work that evoked the difference of concentrations is converted into the potential energy of the hydrostatic pressure. Again, whether the potential energy created by this work is preserved or not depends exclusively on the conditions of the system, that is, on its structure, in the given case on the structure of the semipermeable partition. If the structure of this partition is itself an equilibrium one, that is, does not change without a change of the conditions from without, then the potential energy is preserved; if, however, the structure is non-equilibrium, then the potential energy is not preserved, but is again equalised.
And so, we see that the conclusion to which the first law of thermodynamics and the results of Rubner’s experiments compel us, and according to which the work performed within the organism by the chemical energy of the food is not preserved in the form of potential energy, but is constantly equalised at once and given off in the form of heat—that this conclusion contains within it an assertion concerning the structure of living systems. It asserts that the structures of living systems are not equilibrium ones, that for their preservation, that is, the preservation of the conditions of the system, it is necessary constantly to renew them, that is, constantly to expend work. Thus, the chemical energy of the food is employed in the organism for the creation of the free energy of the structure—the free structural energy [свободная структурная энергия — svobodnaia strukturnaia energiia]—for the building-up, renewal, and preservation of this structure, and is not directly converted into work. The result of Rubner’s experiment in itself admits, of course, of another interpretation as well, namely, that every work of the organism appears anew in the quality of heat of friction, and that consequently the organism represents a machine running idle. This, however, contradicts the fact that for the preservation of the organism, of “the component parts of this machine,” and of its capacity for work, the chemical energy of the nutritive substances and their exchange are necessary. Whereas the chemical energy of fuel is not at all required for the preservation of the component parts of any machine, in living organisms it is necessary for the preservation of the capacity for work of the structure of the systemic forces. The work itself, however, is performed directly at the expense of the structural energy [структурная энергия — strukturnaia energiia]. The potential energy of the structure performs work upon its equalisation. In living systems only systemic forces [системные силы — sistemnye sily] are the source of external work. When a muscle performs work, this occurs not directly at the expense of any chemical processes supplying energy, but at the expense of the potential energy of its structure; the chemical energy, however, serves for the maintenance and restoration of the structural energy. The constant direct conversion of chemical energy into the free energy of the structure is therefore a necessary condition for the preservation of the capacity for work of living systems; it is an inevitable consequence of the results of Rubner’s experiments and contains within it the conclusion that living structures are not in equilibrium, that is, that under the given conditions of the system they would equalise themselves without this constant new formation and maintenance. Such a direct conversion of the chemical energy of the food into the free energy of structures is of course not work in the physical sense; in physiology this process is often called static work. If we, for example, hold an outstretched arm in a horizontal position, then our body in this position performs no work in the sense of thermodynamics; by the mere preservation of this position one can neither stretch a spring, nor raise a weight, nor compress a gas. But for the maintenance of this position of the arm constant energetic processes are necessary, which maintain the free energy of the structure in the given position, since the latter is not an equilibrium one under the given conditions of the system of the articulations, and so forth: upon the switching-off or diminution of this conversion of chemical energy into the free energy of the structure the arm immediately drops. For the maintenance, however, of the horizontal position of the cross-beams of a cross driven into the ground, there is no need of such a constant conversion of chemical or other energy into the free energy of the structure, that is, static work is not needed. This occurs not because the system of the cross does not possess free energy, but because under the given systemic conditions, with the given structure of the wood and of the fastenings, this system is in equilibrium.
With just such static work we have to do when the concentration of some salts within a cell is higher than in the surrounding environment. This state is maintained by the semipermeable structure of the membrane, which is non-equilibrium and, consequently, for its preservation requires a constant conversion of chemical energy into the free energy of the structure of the membrane. For the maintenance of such a difference of concentrations in cells enclosed in a membrane of collodion or parchment, or in Traube’s cells, there is no need of these conversions of chemical energy into the free energy of the structure, since, although the system also possesses free energy, it is yet, under the given conditions, with the given structure of the walls, in equilibrium.
How, then, does the matter stand in such an osmotic cell with external work, that is, with work in the sense of thermodynamics? Is it really the case here too that our proposition holds, that external work is performed only by systemic forces?
With respect to muscular work this was self-evident on the basis of all that has been said above. With respect, however, to the work of an osmotic cell the matter at first glance stands otherwise. We know, after all, that an osmotic cell can perform work even without a change of the structural energy of the membrane, without the systemic forces performing work. Let us imagine, for example, a partition fitting closely against the walls of a cylindrical vessel, but movable. Beneath it there is in the vessel a concentrated aqueous solution of some salt, and above it—distilled water or the same solution, but of lesser concentration. If the partition is loaded with a weight smaller than the osmotic pressure acting upon it, then this system will perform work: the weight will rise until its heaviness becomes equal to the osmotic pressure acting upon the partition, and until equilibrium thus sets in. In this case we have at the outset a system in a non-equilibrium state, performing work at the expense of its free energy of the difference of concentrations. The structure of the system, the internal state of the walls, in particular of the semipermeable, movable partition, remains unchanged in the process. The possibility of such work, however, depends on the presence of a non-equilibrium system, which must first be created by external work directed against the system, as in a wound-up machine. On the other hand, in so far as the structure of the system, the internal state of its walls, does not change (see also the example on p. 25), this work leads to the formation of the potential energy of the raised weight, which is already preserved. We have already shown above that precisely this preservation of potential energy is, on the basis of Rubner’s experiments and the first law of thermodynamics, excluded for a living system. Consequently, into such work the chemical energy of the food cannot be converted. This energy can only serve for the maintenance and creation of the internal structures of the living system, which themselves are not in equilibrium.
We arrive at the general proposition that the structures of living matter possess free energy, which can diminish under the conditions existing at each given moment, that is, that these are non-equilibrium structures, and that in living systems all work within them or directed against their change can be performed only at the expense of this structural energy, that is, by systemic forces. This proposition we shall henceforth call the principle of the work of systemic forces [принцип работы системных сил — printsip raboty sistemnykh sil]. As we have seen, it contains within it the principle of stable non-equilibrium of living systems. On the other hand, the latter contains within it the principle of the work of systemic forces. Both principles signify one and the same thing, but in a different form and from different sides. The cognition of the significance of these principles in both forms is exceedingly essential for the understanding of a whole series of processes.
One of the most graphic and essential differences between living systems and machines becomes clear precisely by means of the second formulation of our general law of motion of living matter. In working machines of any kind work is performed not at the expense of a change of the states, of the structure of the component parts of the machine, and the systemic forces perform work only in so far as the parts of the machine are set in motion by a source of energy external with respect to them. In every machine the internal state of its parts remains practically unchanged (if one abstracts from friction, wear, and the like); the component parts of the machine serve only for the transformation into work of some kind of energy, for example chemical, thermal, electrical. In living systems, however, what is transformed into work is the energy of the internal build, of the structure of living matter. The energy of nutritive substances taken in from without is not transformed into work by means of the structure of living matter and its parts, but goes to the renewal and maintenance of the internal build of this structure and of its component parts. Consequently, living systems are not working machines in the usual sense of this word. The chemical energy of the food taken in by them is not converted in them directly into work. This is precisely the conclusion to which we arrive in applying the first law of thermodynamics to the results of experiments obtained on living beings.
But living beings are not heat-engines either, that is, the chemical energy of the food is not converted in them directly into heat. This is shown us by the application of the second law of thermodynamics to living beings. In view of the fact that this question is expounded in almost all textbooks of physiology, we shall here touch upon it only briefly.
From the second law of thermodynamics it follows that a quantity of heat at the absolute temperature can be only partly converted into work, a part of the heat being thereby inevitably obliged to pass into heat of lower temperature . The useful actionПолезное действие (“useful action”) is what is today standardly termed thermodynamic efficiency — the ratio of the heat converted into work to the total heat supplied., that is, the ratio of the quantity of heat converted into work to the whole quantity of heat,
is equal in this, in the ideal case (with reversible processes), to
But experience shows us that the useful action of a working muscle is equal to approximately 30%. If, consequently, in the muscle the chemical energy were converted into work only after a preliminary conversion into heat, then would be equal to the temperature of the body, that is, 310°, and consequently the muscle would heat up to approximately 170°, which is impossible in view of the ensuing coagulation of the proteins.
From this it was concluded that the living organism works not at the expense of heat, that is, in this senseThe source reads «в этом смысл?», where the question mark and truncated «смысл» mark a garbled spot in the text; read here as «в этом смысле» (“in this sense”). is not a thermodynamic but a chemodynamic machine. This is supposed to signify that the chemical energy in the organism is directly converted into work. Such a relation approximately obtains, for example, in the case of a machine set in motion by accumulators. But we have shown above that this conclusion too is incorrect, that the chemical energy in the organism is likewise not converted directly, by means of any transformers, into work. Living systems are neither thermodynamic nor chemodynamic machines. They are in general not machines in the usual sense of this word; they are subject to special, peculiar laws, by which they differ from machines and non-living systems. The laws of thermodynamics retain in this their significance, but are manifested in a quite different form and cannot be applied to living systems directly, without taking into account these special laws and the special state and build of living matter. Only in the case where we shall constantly bear in mind these special laws, the special state and build of living systems, shall we be able to understand the process of metabolism and shall secure ourselves against errors in the application of the laws of thermodynamics.
Chapter 3: The Contradiction between “External” and “Internal” Work in Living Systems. The Principle of Increasing External Work as a Historical Regularity
The law of motion of living matter, expressed in the preceding sections in a twofold form, means, as we see, that work in the physical understanding of the word, or, as we shall designate it for brevity, “external work” [внешняя работа — vneshniaia rabota], can be produced only at the expense of the free structural energy of living matter. At the same time, however, it follows from this law that all the energy taken in from without, liberated in the organism in consequence of processes of equalisation and converted into work, goes to the maintenance of the non-equilibrium state, that is, of the capacity for work of the organism, to the construction and maintenance of structures; in other words, it is transformed into free structural energy. If, in contrast to “external work,” we designate this constant transformation of free energy into structural energy (called above “static work”) as “internal work” [внутренняя работа — vnutrenniaia rabota], then our law will essentially mean that living matter produces only internal work. External work in the sense of physics, in the sense of the work of a machine, thus stands in contradiction with the general laws of motion of living matter. Since external work can take place only at the expense of the structural energy of living matter, every external work must be accompanied by a partial or complete destruction of this structure. But since, by the law of motion of living matter, all the structural energy and all the energy taken in from without serves for the maintenance and restoration of this structure, that is, is transformed into internal work, then the result of this internal work is destroyed by the external work. By virtue of our general law of motion and the considerations set forth in the preceding section, the structural energy converted into external work can no longer be employed for internal work, that is, for the maintenance and preservation of non-equilibrium structures, and consequently of the capacity for work of the organism. Such energy is already finally lost for internal work and can be restored only at the expense of energy taken in from the surrounding environment. And this will always occur, by our law of motion, provided only that it is possible under the given conditions. Consequently, if our law of motion is valid, then external work can be produced by living systems only when these systems are in some manner disturbed by external actions. These disturbances must hinder the conversion of free energy into internal work, that is, the maintenance of the structure at the expense of the free energy of living matter, and thereby liberate this energy for external work. Such external actions, which disturb the living structure and its form of motion or temporarily switch it off and thereby render “external work” possible, we call stimulations. Further, in the chapter on stimulation and excitation, we shall see that precisely in this consists the role of stimulations. Here it is important for us only to establish that external work is possible only in consequence of a temporary disturbance of the living structure or of its form of motion, and that external and internal work stand in contradiction with one another, since the former is accompanied by the destruction of that which was created by means of the latter.
But this contradiction between external and internal work is of necessity given in the very arising and existence of living matter. It is inseparable from the essence of living matter and its laws of motion. If living matter arises or has arisen anywhere, then it cannot constantly exist as such without producing external work. Therefore irritability, even if in the very weakest degree, is a general property of living matter. This is the result of the laws of motion of living matter. We see that, on the one hand, for the preservation of the non-equilibrium of the work-capable structure of living systems, the latter must constantly produce internal work, for which there are necessary the taking-in of energy from the surrounding environment and its transformation into structural energy. This transformation of the energy of the food taken in into structural energy occurs, it is true, at the expense of the free energy already given in the living structure itself; but its taking-in from without and its conveyance to the individual particles within the organism cannot take place without the expenditure of external work. For, as we see, the source of the capacity for work of living systems is given by the very structure of living systems, by the very structure of their parts, and does not lie outside the system, as in the case of dynamic equilibrium—for example, in the case of a waterfall or a flame. The chemical or solar energy taken in by animals and plants from the surrounding environment cannot be taken in and converted into the energy of the work-capable structure of the living system without this system’s producing external work. This is impossible even in the case where we take care that these sources of energy are constantly and in sufficient quantity conveyed to the organism. Let us explain this by a few examples.
When we consider the most primitive representatives of the animal world—for example the amoeba—it becomes clear to us that for the taking-in of food into the organism a certain degree of external work of change of form is necessary. This work of change of form is necessary even in the case where we cultivate the animal under the conditions of an experiment and constantly take care that the corresponding nutritive substances are present in the culture medium. Under natural conditions the latter does not occur, and therefore a new necessary condition for the taking-in of food from the surrounding environment is added: displacement, the cell’s own movement, which in the given case is accomplished, as is known, by means of a constant change of form, the attachment and drawing-up of the body, that is, again accompanied by external work. The work contained in our general law of motion against the equilibrium expected upon a change of the environment (that is, adaptation [приспособление — prisposoblenie]) thus, in the given case, presupposes the work of locomotion as a necessary condition. The conversion of the chemical energy of the taken-in particles of food into the structural energy of the various parts of the organism in turn presupposes of necessity the presence of certain directed currents within the organism, that is, again work in the sense of physics. We see that a certain, even if minimal, external work is necessary for the maintenance and existence of living matter and its motion.
As evidently follows from our example, the presence of external work is necessary in all those cases where the chemical energy necessary for the maintenance of the structure of living matter is taken in from the surrounding environment directly in the form of organic nutritive substances. But this is not always the case: for example green plants, for which the external source of energy is sunlight, convert this energy into chemical energy within the organism by means of chlorophyll. Here the necessity of external work in the form of changes of form and of locomotion for the taking-in of food falls away. Before going further, however, we must already here remark that, contrary to a very widespread opinion, neither the plant cell, nor even the chloroplast alone, is by any means a simple transformer with a determinate structure that is in equilibrium and changes only in so far as light acts upon it. In actuality we here have precisely the reverse: the chloroplast serves as a photochemical transformer only so long as its structure is in non-equilibrium and is maintained, or newly formed, by means of a constant conversion of the chemical energy of the organic substances in the cell—substances synthesised by means of the chloroplast itself—into the free energy of the structure of the chloroplast, that is, by means of continuous internal work. Without the energy-supplying process of respiration the chloroplast perishes; the chloroplast isolated from the cell is incapable of transforming solar energy into chemical energy. The extent to which photosynthesis depends on the structure of the rest of the protoplasm, and the extent to which it is maintained at the expense of the structural energy of the protoplasm, is shown by the recent interesting experiments of Lyubimenko.[Author’s note:] Lyubimenko and Shcheglov, Planta, vol. 18, no. 1/2, p. 383. Consequently, in this case too, the case of the utilisation of solar energy, our laws of motion of living matter are in no degree affected. With this manner of nutrition there falls away only the necessity of taking in organic nutritive substances directly from the surrounding environment, and consequently the necessity of that work which is expended upon this, that is, of change of form and of locomotion. Indeed, we see that these two kinds of external work, encountered in all animals, are absent in plants. Consequently, the moment of external work in plants recedes into the background by comparison with internal work. Does this mean that external work is here altogether absent, or that it can even merely be absent? By no means. There still remains the necessity of protoplasmic currents within the cell for the distribution of organic substances within it, the necessity of work upon changes of external conditions—for example, upon a change of illumination. This latter form of work need not necessarily be expressed in the form of a change of place; it may be expressed by means of a change of the position of a leaf or of the position of the chlorophyll grains, of their distribution or of the rate of their displacement within the cell. But each of these forms of adaptation, that is, of work proceeding from our general laws of motion against the equilibrium expected upon a changed environment, is work in the physical sense of the word and is consequently produced at the expense of the energy of the living structure.
Thus we see that the contradiction between external and internal work appears already with the arising of the form of motion of living matter, that is, with the arising of this matter itself. It is conditioned, on the one hand, by the fact that living systems must take in from without the energy needed for the maintenance of their capacity for work, must adapt themselves to external conditions, and, on the other hand, by the fact that external energy within the living system is not transformed directly into physical work. It is entirely obvious that the role and significance of external work in relation to the fundamental form of motion of living matter—internal work—may be, and in various living beings actually are, very different; they depend, on the one hand, on the structure of the living being, and on the other, on the surrounding environment, that is, on its sources of energy and on the changes of the latter. From the general laws of motion of living matter it naturally follows that every living being must possess certain work-capable structures, at the expense of the equalisation of which, that is, of their partial and temporary destruction, external work can be produced for the taking-in of energy from without and for adaptation to the fluctuations of the latter. In this sense every living being is more or less adapted to the surrounding environment. Which sources of energy can be utilised by means of these structures, or by means of external work at the expense of the energy of these structures, and upon which changes of the sources of energy external work for adaptation can still be produced—this depends, of course, on the structures themselves. We have already seen, in the cases adduced above, that, for example, green algae can utilise the energy of sunlight for the creation of chemical energy within the organism, while unicellular animal organisms, under the same conditions, are incapable of utilising this source of energy. We have also seen that this capacity to produce chemical energy within the organism has an essential significance for the determination of the role of external work. We know of bacteria which are in a position to utilise the energy of the oxidation of iron or of the nitrogen of inorganic salts for the synthesis of organic substances within the organism. Thus, in these organisms there falls away the external work necessary for adaptation to fluctuations of light energy and for the taking-in of organic substances, that is, that work which, under the same conditions, other organisms must produce. Probably here too the external work necessary for the taking-in of inorganic substances and their distribution within the cell is smaller, since it is possible that to a great degree there is utilised the work of osmotic forces and the diffusion of substances in the soil, which require no external work at the expense of the structural energy of the organism. Besides this, there are known to us animal organisms—parasites—which, it is true, take in organic substances from without, but in which the external work expended upon this is reduced to a minimum, since they live within other animal or plant organisms, which produce this work for them. Thus we see that the degree and significance of external work may be very different, depending on the structure of the organism and on the surrounding environment. Theoretically one may here admit any possibility, and nature here displays such a diversity as leaves behind it every fantasy.
Can one not, however, establish some regularity in this diversity? Since we here have before us all the forms with all their diverse structures and degrees of external work, it is evident that if there is a regularity here at all, then it must be sought in the history of the development of these forms: it must be a historical regularity [историческая закономерность — istoricheskaia zakonomernost′]. Thus, we must here proceed from the doctrine of evolution and its laws, which has at present been definitively confirmed by experimental means and serves as an unshakable foundation for theoretical biology—a foundation upon which further theoretical constructions can be grounded.
Already in the first chapter, where the subject and method of theoretical biology were discussed, we made clear that evolutionary theory asserts the following: living beings in their contemporary diversity arose neither simultaneously nor independently of one another, but have their own history. They descended from one another, possess a common genealogical tree, and therefore stand in a more or less close “blood kinship” with one another. The origin of the various species by means of selection rests upon the following properties of all living beings: upon reproduction, variability, and heredity. On the basis of these properties the origin of species finds its full natural-scientific explanation. To avoid all misunderstandings and tautologies, we shall not separate the last two properties from one another and shall speak only of hereditary variability, that is, of mutations, according to the usual terminology. We then have to do with only two properties: reproduction and mutability. Evolutionary theory gives the following grounding of the arising of new forms: the reproduction of living beings leads with inevitability to the dying-out of the majority of individuals before they can reproduce anew. If organisms did not possess the property of mutability, then new species could not arise, and which individuals survive and reproduce further would be left wholly to chance, since they are all alike, with equal probability fall into more or less favourable conditions, and, possessing one and the same structure, can utilise only the same sources of energy within the same limits of their fluctuations. But since organisms possess mutability, their internal structure is subject in individual specimens to changes, so that the probability of survival is not the same for all individuals. In the first place, various organisms can, owing to even minimal differences in their structure, adapt themselves in various degrees to the fluctuations of external conditions, and, in the second place, the probability of falling into more or less favourable conditions becomes unequal. If, for example, a given animal becomes more mobile, or the seeds of some plant, owing to their structure, are more easily borne by the wind, then for them the probability of falling into more (but also into less) favourable conditions naturally increases. Consequently, those individuals will survive with greater probability which, in consequence of hereditary variations of their structure, can either adapt themselves to greater fluctuations of the environment or with greater probability fall into more favourable conditions. By environment we understand, of course, not only inorganic nature, but also the density and composition of the population formed by other living beings. The point is that, on the one hand, there sets in among them a competition for the limited sources of energy of inorganic nature, such as water, light, soil, for nutritive substances, and so forth, while, on the other hand, they themselves serve as food for one another and mutually exterminate one another. Which new structures arise, which structures increase the probability of survival in this “struggle for existence,” when there is a greater probability of their appearance, and whether their arising stands in any connection with the external conditions of existence, as well as with survival and reproduction—of all this evolutionary theory says nothing. Moreover, the arising of hereditary variations is regarded simply as a property of living organisms, and in contemporary biology there is denied any regularity concerning the forms and frequency of variations, and consequently any law-governed dependence upon the survival or dying-out and upon the reproduction of organisms. Which variations appear and when they arise is consequently a matter of chance. In the chapter on evolution we shall see that this view of the contemporary doctrine of evolution is conditioned by an insufficient knowledge of the general laws of motion of living matter, and that the application of these laws in this domain too permits us to penetrate more deeply into the essence of phenomena and to establish certain regularities. Here we shall only show that, on the basis of the contemporary evolutionary doctrine and our general laws of motion of living matter, we arrive at a regularity in the history of the development of organisms, concerning the contradiction between external and internal work.
Although the appearance of hereditary variations of structure is purely accidental, the survival of one or another individual occurs already not with equal probability. As we have seen, those individuals possess the greater probability of surviving which can either adapt themselves to greater fluctuations of the surrounding environment or with greater probability fall into favourable conditions. But we have seen that both the adaptability to changes of the sources of energy in the surrounding environment, taken in with a given structure, and the probability of falling into more favourable conditions (that is, the mobility of organisms) are in general the greater, the more external work can be produced under otherwise equal conditions. Of course, this rule is valid only in the most general outline; this connection is by no means immediate: neither the probability of falling into more favourable conditions nor the adaptability to greater fluctuations of the sources of energy in the surrounding environment is raised, or can be raised, only by means of an increase of external work. Thus, for example, a greater mobility, that is, a greater probability of falling into favourable conditions with unchanged structures serving for the taking-in of energy, may be conditioned not only by an increase of the external capacity for work, that is, by a greater proper mobility of the organism, but also by such a structure as utilises the forces of the surrounding environment, with which the work necessary for locomotion is produced by these external forces. We observe this, for example, in the structure of certain seeds, which are more easily borne by the wind, and such a structure creates for them a greater probability of falling into favourable conditions without any strengthening of external work. On the other hand, the adaptability to fluctuations of the sources of energy may be raised, even with an unchanged environment, not only by means of an increase of external work: for example in plants the utilisation of the energy of sunlight may be raised not only by means of structures permitting external work to be produced—such as the turning of leaves, the displacement of chloroplasts within the cells, and the like—but also without an increase of external work, by means of such structures as, for example, a large surface of the leaves and the like. The same applies also to animal organisms, whereby here too the adaptability to greater fluctuations of the environment may be attained by means of structures which permit, for example, in the taking-in of food, a better utilisation of the osmotic forces of the surrounding environment without an increase of the external work of the organism. Finally, there arise new structures permitting the utilisation of new sources of energy of the surrounding environment, as we have seen in the example of the iron and nitrogen bacteria. The taking-in of these new sources of energy is not necessarily accompanied by an increase of external work; on the contrary, we have seen that in the adduced examples the reverse is the case. Thus, green plants must produce considerably less external work, owing to their capacity to utilise the energy of sunlight for the formation of chemical energy within the organism. Further, we know that parasites and symbionts, which can utilise the work of other organisms or the chemical energy formed by the latter, must produce for this not more but, on the contrary, less external work. By means of these new structures, permitting the utilisation of new external sources of energy for internal work, that is, for the maintenance and creation of the non-equilibrium structures of living matter, the probability of survival in the struggle for existence is likewise increased.
Thus we see that the greater probability of survival in the struggle for existence may be secured not only by means of an increase of external work, and is therefore not always accompanied by such an increase. But parallel with the increasing density of the population, which is the inevitable consequence of the reproduction of organisms, and with the increasing spread of new forms over new territories in consequence of the arising of new structures permitting the utilisation of new sources of energy—parallel with this there inevitably proceeds a rise of the role and significance of external work by comparison with internal. On the one hand, with the increasing density of a population of identical organisms, that is, of organisms utilising identical sources of energy, the probability of survival is the greater, the greater the external work that the given organisms can produce owing to their structures. On the other hand, even upon the arising of structures permitting the utilisation of new sources of energy, and consequently the settlement of new regions hitherto weakly or even not at all settled, the greater probability of survival is secured by these new structures only so long as, in this new environment too, the density of the population, in consequence of reproduction, does not increase to such a degree that survival again becomes possible only by means of structures permitting once more either to increase external work or to utilise new sources of energy, and so forth.
We see that the historical development of the diversity of forms of living systems must have proceeded in two directions: 1) in the direction of an increase of the diversity of simultaneously existing forms with respect to the utilisation of various sources of energy and various external conditions, 2) in the direction of an increase of external work within the limits of one and the same group of forms. One must, however, by no means think that the various groups of forms exist independently of one another, even if they utilise various sources of energy. They coexist in time and in space and therefore constitute the surrounding environment and the source of energy for one another. The chemical energy formed by plants within the organism by means of sunlight is utilised by animals as a source of energy. The chemical energy formed by the host-organism is utilised by the parasite, whereby harm is done to the host. One species of animals lives at the expense of another, and so forth. In this struggle for existence external work, evidently, acquires an ever greater significance for the probability of survival. The probability of survival is raised not only by the fact that, within the limits of identical forms or of forms standing in close kinship, the structures, under otherwise equal conditions, afford the possibility of producing greater external work, but also by the fact that the degree of external work in one form is higher than in another.
Thus, we arrive at the following conclusion: in the course of the arising of the diversity of forms of living beings, the role of external work becomes ever more important, in consequence of which the latter must increase. This regularity is not a direct but a historical one, realising itself of necessity through the single and accidental. In particular, from the considerations adduced above there follows this: among identical or kindred forms which, for internal work—that is, for the maintenance and creation of their non-equilibrium structures—make use of identical sources of energy in one and the same locality, the more ancient forms possess less developed structures for external work than the later ones. Or, what means the same: the newly arising forms, which can utilise new sources of energy and a new surrounding environment, at first possess a lesser capacity for external work; the latter falls in them to a minimum, in order then to rise anew within the limits of that same group of forms. An example may be afforded by plants utilising light energy, by parasites, by nitrogen and thermophilic bacteria, and so forth. Finally, among all simultaneously existing forms the maximum of external work increases with time.
Here it is fitting to say a few words about the character of this regularity, the more so since such historical regularities, which speak of a directedness in the development of species, very often arouse among biologists a fright that is understandable, though not always well-founded, and give occasion to a whole series of misunderstandings. The cause of this, as we have already said in the first chapter, is that such “tendencies” in the direction of development can very often lead to vitalistic conceptions and still more often proceed from them. The so-called “ladder of living beings” of Lamarck, the various theories of orthogenesis—all of them have beneath them a vitalistic foundation. This is conditioned by the fact that the vitalists themselves cannot picture to themselves any regularity other than the immediate and essentially externally causal one. When they observe, or think that they observe, some regularity in development, they consider it necessary to assume for it too an immediate, externally acting cause lying outside, some purposive entelechy. Then to the causal regularity there is opposed a so-called final, goal-striving regularity. This opposition is fundamentally incorrect, since from the fact that a regularity cannot be reduced without remainder to externally causal relations there by no means follows the necessity of a final regularity, of purposiveness, that is, of a teleological moment. Thus, such an opposition in the sense of tertium non datur is incorrect. There exist historical regularities which are not exhausted without remainder by externally causal relations and which nonetheless have nothing in common with purposiveness, entelechy, or teleology. But the opposition of final and causal regularities in the sense that the former is allegedly a different explanation, not based on causal connection, is likewise incorrect, since when the vitalists consider themselves obliged to have recourse to a final regularity, they nonetheless cannot in so doing free themselves from the conception of a purely causal connection as the only possible exhaustive regularity, which is why they consider it necessary to have recourse to a cause lying outside in the form of an entelechy, thus reducing the given regularity in the end once again to a purely externally causal one. Of historical regularities based on the laws of motion of living matter themselves, with their inevitable interaction with the surrounding environment, they know nothing. Unfortunately, with some materialists the matter often stands little better: although they speak of historical regularities, they reject them as idealistic, and therefore do not see them, thus throwing out the baby with the bathwater.
Therefore the question is fitting: when have we the right to speak of a historical regularity, and in what does it consist? In what is its difference from the purely causal one? First of all, it is clear that a law-governed change in time is not an essential mark of a historical regularity. The motion of the earth around the sun and its position in space depend strictly on time, and we can predict its position with great accuracy for days, years, and tens of years ahead. Nonetheless, we here evidently do not have to do with a historical regularity. This regularity in time is unequivocally determined by the acting causes of the attraction of masses and the position of the heavenly bodies relative to one another. The same takes place also in the fall of a body in a vacuum, in so far as the resistance of the air can be neglected. Here too we have, expressed by the laws of fall, an exact dependence of the position and velocity of the falling body upon time—a dependence likewise deducible without remainder from the mutual attraction and the spatial relations. But how stands the matter, for example, with the following regularity: every body whose specific gravity is greater than that of the air will sooner or later fall, despite the wind and the resistance of the air, and independently of the accidental directions and force of the wind and of the position of the body relative to the earth? Here we have before us a regularity realising itself of necessity in time through the particular and the accidental. A flying leaf may be raised upward, in the next moment fall down again, and so forth; every subsequent moment of the motion is to a considerable degree independent of the preceding one, cannot be determined in advance, and nonetheless it can be predicted that in time the leaf will fall to the earth. Is this regularity perhaps a historical one? By no means. The point is that this regularity too is conditioned by an acting cause that remains unchanged in time, namely the attraction of the earth. We have an unchanging field of attraction of the earth, constantly acting in one direction—toward the centre of the earth—and therefore it must be regarded as acting independently of the movements of the air, as a cause lying outside. But in order that there should be a historical regularity, it is not sufficient that the acting cause—in the given case, for example, the field of attraction—should itself change in time, since this does not change the matter essentially, but only complicates the causal relations. In order that the field of attraction might be regarded not as a cause lying outside, there must exist a dependence between the movements of the air and the changes of this field in time. But then there arises the question: how, then, is it possible that the regularity of falling to the earth realises itself of necessity, if the movements of the air are not themselves subject to a determinate regularity, but are accidental? For if there exists a law-governed dependence between the changes of the movements of the air and the changes of the field of attraction, then the latter must likewise bear an accidental character, and the fall of the leaf will not realise itself of necessity.
From a purely externally causal standpoint this is incomprehensible. For if one variable—for example the direction of the movement of the air—is some unequivocal function of time, and another variable—in the given case the direction of the field of gravity—is likewise a function of time, then, given a law-governed connection between the two, the character of the changes of one variable is of necessity given by the character of the changes of the other. For the changes of one variable in time can proceed according to a determinate regularity (for example, monotonically increasing or periodically), despite the accidental character of the changes in time of the other variable, only if both are independent of one another. Such a standpoint is, however, correct only in the case where we consider that the mutual dependence of the variables is realised only under the following conditions: 1) when the law-governed dependence of the two variables is mutual and unequivocal, that is, the changes of the one determine the changes of the other and vice versa, 2) when this dependence does not itself change in time, 3) when time as an independent variable does not play a special role in consequence of some factor of inertia, that is, when either the arising of some change is just as probable in time as its disappearance, or this factor of inertia may be neglected. But all these three conditions will not be fulfilled if certain essential laws of motion of the system overlie this law-governed dependence between the two variables, and nonetheless such a dependence will exist. For the first point let us adduce the following examples: every change of temperature in the surrounding environment naturally influences the temperature of the body, and yet the body temperature of warm-blooded animals exhibits law-governed, periodic fluctuations in the course of the day, despite the accidental changes of the external temperature; a rise of temperature up to a determinate limit leads to an acceleration of cell division, but not the reverse; the injury of a cell leads to division and regeneration, but not the reverse. In general, wherever we have specific, proper laws of motion of the system, the unequivocal mutual dependence falls away. For the second point let us point out that the law-governed dependence between two variables may change in time, if the internal laws of motion themselves change in consequence of a change of the internal structures, but not in consequence of new causes acting from without; the latter would lead only to a new form of those same laws of motion. Such changes are abrupt and are bound up with essential qualitative changes. From biology one may adduce mutations as an example of them. For the third point let us point to the well-known concept of irreversibility, or hysteresis. All these phenomena of after-effect are based on the internal structure and on the proper laws of motion of the system, and not on a cause acting from without. But whereas in hysteresis this factor of inertia, this factor of after-effect, plays an insignificant role by comparison with the fundamental forms of motion, does not overlie them, and is, so to speak, only a correction to the fundamental laws of motion, in biology, and specifically in the question that interests us, the matter stands fundamentally otherwise: here this factor of inertia is an essential, fundamental property of living matter itself—we speak of heredity.
Thus we see that we have a historical regularity when the system moves in time on the basis of the internal laws of motion proper to it, and not only by virtue of causes acting upon it from without, and when, in consequence of these laws of motion of the system, there inevitably sets in a law-governed change in time of certain of its properties, of certain variables, although the changes of other properties, causally connected with the given variable, occur accidentally.
From what has been said it follows that a historical regularity can be understood only on the basis of the general laws of motion proper to the given state and the given structure of matter, and that only from these does this regularity follow. Chronological dependence in time, even if it is law-governed, is still far from being a historical regularity. A strict temporal dependence is characteristic precisely of causal, dynamic laws, whereas for historical regularities it is characteristic precisely that only the inevitable direction of the changes in time can be indicated, but not a strict temporal dependence. It is precisely for this reason that a historical regularity cannot be found by means of a pure generalisation of empirical regularities without a far-reaching theoretical analysis and a knowledge of the general laws of motion of the given matter.
After these brief considerations concerning the character of the historical regularity, which we considered necessary in order to avoid misunderstandings, and especially in order to avoid any occasion for confusion with vitalistic tendencies, we may return to the regularity put forward by us—to the principle of increasing external work. According to this principle, both contradictory tendencies of internal and external work are given already in the very arising of living matter by virtue of the laws of motion proper to it, and external work increases of necessity with the lapse of time, that is, in the history of living beings on earth. We have shown that this is a necessary consequence of the general laws of motion of living matter, and that the increase of the significance of external work occurs with inevitability, even if the other variables which condition this increase, and on which it therefore directly depends—namely the appearance and the form of hereditary variations—change purely accidentally.
In the chapter on evolution we shall deal specially with the question of the mutual dependence of external work, physiological adaptability, and the arising of hereditary variations, and we shall there set forth a theory according to which the appearance of such variations is not independent of these variables. But the increase of the significance of external work, and consequently of the contradiction between external and internal work, is nonetheless preserved independently of what this immediate dependence is and of whether it is subject to chance or not.
Let us now see to what extent our principle is confirmed by experience. We do not, it is true, have at our disposal sufficiently systematic measurements of the degree of external work and of its relation to internal work to use them directly as a proof, but we can nonetheless adduce two extreme cases: one in the unicellular organisms, the other in human beings. The relation of external work to internal work in the protists has been studied in detail by Ludwig on paramecia. He measured the consumption of oxygen and calculated how much work the organism produces for its locomotion. The various methods of measurement and calculation led to sufficiently concordant results, according to which less than 1% of the whole energy obtained by means of respiration is expended upon the work of movement. More than 99%, consequently, goes to internal work and to external work within the organism. In human beings, on the other hand, the metabolism in the state of repose amounts, as is known, in general to less than 2000 large calories per day, while in heavy physical labour it rises to 4000–5000 calories. Here, consequently, we may reckon that on the average 50% of the whole energy goes to external work in the sense of work directed against the surrounding environment, while only 50% goes to internal work and to external work within the organism, such as the work of the heart, the osmotic work of the kidneys, and the like. With this it is highly probable that the part falling to external work within the organism in man and in mammals is also relatively considerably higher than in the infusoria. Although we do not have sufficient quantitative data concerning the intermediate steps, we nonetheless have at our disposal very important indications, clearly speaking in favour of the growth of the significance of external work. External work directed against the surrounding environment is produced by animals by means of contractile elements. The tissue specially differentiated for external work is muscular tissue, especially the striated muscles. Smooth musculature is an intermediate step; in part it too already serves for external work, but chiefly for the maintenance of determinate tensions at the expense of internal work. Correspondingly, we see that in the most ancient forms of the unicellular organisms we either do not meet with these elements at all, or else in the more complex, later forms there appear at best simple contractile elements, such as the contractile vacuole of a number of unicellular organisms, the contractile elements in Vorticella, and the like. In the coelenterates, too, true musculature is still rarely encountered. They possess a so-called muscular epithelium, the contractile elements of which—for example in the hydra—already group themselves into a thin layer; by our observations in polarised light this layer exhibits, in the stretched state, double refraction, which disappears upon contraction; in the worms, smooth musculature appears and becomes predominant, while the predominance of striated musculature appears for the first time only in the Arthropoda. Finally, in the phylogenetically youngest forms, in the mammals and in man, 50% of the weight of the body falls to striated musculature, that is, to the tissue specially differentiated for external work. These morphological data, showing the law-governed increase of determinate structural elements and the historically law-governed change of this structure with time, confirm, of course, sufficiently clearly our proposition concerning the existence of a historical regularity.
We can, however, find a confirmation of this regularity also from another point of view. Above, in speaking of the principle of the work of systemic forces, we pointed out that one of the most essential differences between a machine and a living system consists in this, that machines produce work not at the expense of the free energy of the structure of their parts—these parts remain unchanged in their structure and serve only as transformers—while in living systems the matter stands precisely the reverse. But if, in the course of the history of development, the significance of external work increases of necessity, then in the structure of the organism this must be reflected also in yet another way: there must increase not only those elements which can produce external work at the expense of their structure, that is, the muscles, but there must also increase those elements which make possible the transformation of internal work into external in the sense of a power-machine. There must, consequently, develop such parts of the system whose internal structure during work almost or entirely does not change, but which are suitable for the transmission of work. Such elements we find in the parts of the skeleton and their structure in the higher animals (arms in the form of levers, articulations, and the like). Whereas in the more ancient forms the hard skeleton fulfils only protective functions in relation to the soft parts and is therefore placed on the outside, we see that, in proportion as the significance of external work and of its source—the structural energy of striated musculature—increases, these parts of the skeleton are displaced ever more inward, in order that the organism might use them as a transformer of internal work into external, and that, in the end, the protective function is limited only to the central nervous system.
Finally, let us touch upon yet another side of the question. We have already said that external work can be called forth only by means of a disturbance of the internal, essential form of motion of living matter, and that these disturbances are called forth by external actions—stimulations. But if the significance of external work in the course of development inevitably increases, then this must naturally be accompanied by an easier disturbability, by a facilitated access of external disturbances, that is, of stimulations; physiologically this is confirmed by the heightened irritability of striated musculature by comparison with smooth. These data must serve only as an example of the fact that we have at our disposal a great number of facts confirming the historically necessary increase of external work as against internal, and moreover that this principle permits us to explain a series of regularities of structure and function in the course of evolution, without having recourse to vitalistic conceptions, but exclusively on the basis of theory and the general laws of motion of living matter. Above we have already pointed out the untenability of the purely causal, unhistorical manner of thinking, not based on these general laws of motion of living matter, which is the chief error of mechanism in biology.
We see thus that, in consequence of the increase of the role of external work, which stands, as has been said, in contradiction with the motion of living matter and can be produced only by means of the destruction of the results of internal work, that is, at its expense, there arise in living organisms ever more moments characteristic of power-machines. This is one of the chief causes why biology freed itself with such difficulty, and even now has not fully freed itself, from analogies with the power-machine and from mechanistic conceptions, the more so since in man and the higher animals external work already plays a fairly considerable role, while its action is most accessible to observation and investigation. It would, however, be a profound error to identify these elements of the organism, characteristic of power-machines, with real power-machines. They are realised in the living organism likewise only through the laws of motion of living matter and on their basis, so that even in man the muscle is capable of work in the sense of external work only in so far as its non-equilibrium structure is constantly maintained and restored at the expense of the energy of metabolism, that is, at the expense of internal work. It is therefore clear that, parallel with the increase of external work, there must proceed also an increase of the structural energy of living matter, a strengthening of internal work.
In conclusion let us further point out that in all plants and animals external work nonetheless serves more or less directly for the maintenance of internal work—for example, for the obtaining of food or of other sources of energy, for work against equilibrium, and for the maintenance and restoration of work-capable, non-equilibrium structures.
As for man, we have already seen that he makes use of external work consciously, and not exclusively on the basis of the biological laws of motion, and not exclusively for the restoration of his own capacity for work. There, where the contradiction between external and internal work is used consciously, where man creates for himself a working instrument, that is, even the most primitive machine—for example a lever—where he consequently begins to make use of machines which no longer require work for the maintenance of the structural energy of their parts, there, but only there, the biological regularities recede into the background and are overlaid by still higher regularities of a higher unit—society.
Chapter 4: The Problem of “Living Protein”
The general laws of living systems set forth by us in the preceding chapters contain, as we have seen, certain assertions concerning the state and structure of the matter out of which living systems are built. We have seen that these general laws contain, as a necessary presupposition, the conception of such a structure as, under the given conditions, is not in equilibrium, the possibility being excluded of not only stable but also unstable equilibrium. Were we to accept the existence of unstable equilibrium, this would contradict the principle of the work of the structural forces and thereby all the factual data from which this principle was deduced. A genuinely unstable equilibrium would mean that changes in the system could be brought about only by means of slight disturbances, that is, changes in the surrounding environment; further, that every such insignificant change in the surrounding environment evokes a change in the state of the structure of the system; and finally, that any, even the slightest, change of the external environment immediately leads to a state of stable equilibrium, that is, that the system loses its capacity for work. All this evidently contradicts the facts. We have already seen that by means of such a supposition we can explain, for example, a single reaction to a stimulus, but cannot explain either the repeated phenomena of stimulation or adaptation. If, however, we suppose that it is precisely this unstable equilibrium that is constantly restored, then the question arises at the cost of what work this takes place, since unstable equilibrium is, after all, distinguished by a maximum of free energy under the given conditions of the system. We should have to transfer the source of the energy of all work beyond the limits of the system, just as was the case with stable and mobile equilibrium.
If the laws of living systems have as their necessary presupposition the non-equilibrium state and structure of the matter out of which these systems are built, then the question arises, by what this state is conditioned. Up to now we have still left this question open, but on the answer to it depends the whole theory of living matter, that is, the conception we form for ourselves of its constitution and state. Thereby is determined both the entire direction and the interpretation and significance of chemical and physical investigations and of the results obtained by their means. Let us first make clear to ourselves the question itself: by what can the non-equilibrium state, and hence the capacity for work, of some system be conditioned, if this state is to be maintained at the cost of the system itself without any influx of work from without? We have already seen that, in contrast to the working machines known to us, such non-equilibrium can be conditioned only by the structure of the constituent parts out of which the system is built. The non-equilibrium state can be explained by the system’s consisting of several phases with different thermodynamic potential, the sum of the thermodynamic potentials of the separate phases being able to diminish under the given conditions of the system. If these conditions remain unchanged, the sum of the potentials will diminish until it reaches a minimum and equilibrium sets in. Consequently, work against the onset of this equilibrium, taking place, as our principle requires, at the cost of the conditions of the system, is possible only in consequence of changes in the state of those parts of the system that border on its various phases. We may imagine that these constituent parts of the system are themselves capable of work, because they consist of several phases with different thermodynamic potential, but then our conclusion must be applied to these constituent parts as well. Thus we see that the non-equilibrium state (that is, the capacity for work of the system without influx of work from without) which answers to our general laws of motion cannot be conditioned by the presence of several phases with different thermodynamic potentials capable of being equalised under the given conditions of the system. But this compels us to come to the conclusion that in a living system changes of state can take place out of one homogeneous phase without change of conditions at its boundaries, that is, without influences from without. This, however, is possible only on condition that the molecules themselves which form the homogeneous phase are in a state capable of changing even without changes of conditions at the boundaries of the phase; and this in turn means that the molecules themselves do not possess a structure corresponding to the state of equilibrium, that is, to the minimum of free energy under the given conditions. Of course, here too we have before us neither an unstable equilibrium, as for example in explosive substances, nor a metastable state of equilibrium, such as Parnas assumes, for example, for the hypothetical maternal substance in the traumatic formation of ammonia (see below). Against the admission of such states for the molecules of the homogeneous phase speak the same considerations that we developed above with respect to multiphase systems, where, as we have seen, they do not correspond to our general laws.
On the basis of the general laws of motion of living systems we arrive at the conclusion that the theory of living matter must proceed from the following proposition: the non-equilibrium state of living matter and, consequently, its constantly preserved capacity for work are in the last analysis conditioned, as our first principle then requires, by the molecular structure of living matter, while the source of the work performed by living systems is in the last analysis the free energy proper to this molecular structure, to this state of the molecules. Or, correspondingly to the second formulation of our principle: external work can be performed only at the cost of structural energy.
Before showing how far this fundamental proposition corresponds to the factual data, and how far from these data a concrete theory of living matter can be developed, we shall briefly dwell on the history of this question and on its significance in principle. The question whether the properties of living beings are based on a specific state, a specific structure, of some definite compound, that is, of some kind of molecule, or whether we have here to do with a combination and joint action of various reacting compounds, conditioned by a corresponding structure—this question is already rather old. The answer that the majority of biologists gave to it at various times depended chiefly on the state of physical chemistry in the given epoch. Before the period of the flowering of physical chemistry, in particular before the deepening of the doctrine of osmotic phenomena, electrolytes, and membrane potentials, on the one hand, and of the phenomena in boundary surfaces, especially of boundary catalyses and of chemical dynamics, on the other, there prevailed among biologists the conception that the characteristic vital phenomena are based on special properties of the protein [белок — belok] situated in the protoplasm of the living cell. Thus Pflüger speaks of “living protein” [живой белок — zhivoi belok]; in Rubner the conception of a special structure of living protein likewise plays an important role. Let us further mention Oscar Loew, as well as Verworn, who put forward the hypothesis of biogen molecules, and his follower Jensen. Common to them is that they ascribed the fundamental vital phenomena—such as metabolism, growth, irritability—to a special chemical structure of the molecules out of which living substance, protoplasm, is built. They assumed a special lability and chemical activity of this compound and endeavoured to explain these specific properties of the hypothetical compound, or of the hypothetical living protein molecule, by means of its chemical structure, which led to the admission of the existence of a special specific chemical compound present in living cells but absent in dead ones, since, by virtue of its lability, this compound readily undergoes chemical changes and passes over into another compound. There is no need here to enter more deeply into these conceptions; they all suffered from the defect that they could not give a concrete theory of this substance, of its physical and chemical characterisation. For this reason they also could not link up the vital phenomena requiring explanation with concrete physical and chemical properties of the hypothetical compound (even if only theoretically deduced), and consequently could not develop a theory of these vital phenomena on the basis of their conceptions, and thus could not go beyond the bounds of pure speculation. The fundamental cause of this is to be sought in the state of the physical and physico-chemical sciences. At that time it was in general not yet possible to give a physico-chemical theory or characterisation of those special states and properties of molecules or of matter which could explain the specific reactions found by physiology and physiological chemistry, their course, their kinetics, and their direction. This is to be explained, on the one hand, by the fact that the phenomena of metabolism, assimilation, growth, fermentation, and so forth were already known at a time when the foundations of general and physical chemistry, the theory of catalysts, and the like were not yet worked out. On the other hand, at that time it was impossible to form for oneself a concrete conception of the state of living matter, since the theory of the structure of matter in general was developed by modern physics only in the most recent times.
The rapid development of general and physical chemistry, which began in the nineties of the last century, led to the striving to explain the various phenomena of metabolism, stimulation, and so forth by means of these sciences. In the preface to the third edition of his “Foundations of General Chemistry,” W. Ostwald writes in 1899 concerning the significance of the development of general chemistry for the other sciences: “The scientific significance of the new ideas can no longer be the subject of serious dispute; it may be recognised by its fruits. But after this progress has made itself felt in general chemistry, there arises a new, extraordinarily many-sided task—to apply the auxiliary methods thus obtained in all those fields whose development stands under the influence of chemistry. On the basis of the new ideas, not only are chemical technology or analytical chemistry being transformed or awaiting transformation, but the more remote fields of science as well, first and foremost physiology, stand before tasks of development and transformation, fraught with consequences, called forth by the application to them of the achievements of general or rational chemistry.” Ostwald’s prognosis was justified: the application of physical chemistry to physiology and biology led to a “development and transformation, fraught with consequences,” of these sciences. For the problem of the theory of living matter that interests us here, this development was of decisive significance: its result was the final interment of living protein, of living matter. The answer that the modern physical chemistry of protoplasm, or of living organised matter, gives to the question posed at the beginning of this chapter runs: living protein does not exist. The properties of living matter are based not on a special chemical compound or on a special state of the molecules, but are explained by the multiphase character of protoplasm and the boundary forces acting at the boundaries of these phases, or by the structure of the membranes separating the latter from one another. This opinion is expressed especially sharply in the following sentence of Höber: “Proteins, carbohydrates, fats—as whatever substances we see them upon entry into the organism—are used as such within it as well; there is no other protein than the one we see in the test-tube; just as there exists no dead, stable protein alongside a living, labile one, so there is no dead and living sugar or dead and living fat. The grounds for the great and constant capacity of living protoplasm for reaction are quite different. They lie in the interaction of all the separate substances in their definite quantitative proportions. In this interaction lies the essence of the problem of metabolism.”
Thus we see that the result of the “fraught-with-consequences” application of physical chemistry to biology, especially to the study of protoplasm, is precisely the opposite of that to which we have come on the basis of our general laws of motion of living matter. It is therefore understandable that the central problem of the modern physico-chemical direction in biology is the problem of permeability and of boundary forces in general. If we seek the source of the capacity for work of living matter, of protoplasm, not in the very state of the molecular structure of definite molecules, but in the different thermodynamic potential of different phases, then of course the problem lies in the properties of the boundary surfaces and in the change of the boundary forces in time, which depend exclusively on the state of the phases meeting at the boundary surface and change only in so far as the phases themselves change. But a separate phase cannot change without a change of the boundary conditions, that is, of the properties of the boundary surfaces, since the non-equilibrium state here could exist only in the case where it were conditioned by the molecular structure of one of the components of this phase. Therefore the modern physico-chemical direction in biology seeks the resolution of the question in changes of the composition of separate phases, of the concentration of separate reacting components, ions, and so forth. But thereby, as the moment that disturbs or shifts the equilibrium, there is introduced as a necessary condition the influx of various substances, that is, there takes place a complete renunciation of the principle of the preservation of the non-equilibrium state which we recognised at the outset. We have before us a constant disturbance of the existing equilibrium in consequence of the influx of substances and energy. The metabolism of the living cell, which is itself to be explained, must explain everything, instead of being itself explained by means of the properties of living matter, its state and structure.
Thus, if before the development of physical chemistry it was impossible to create a theory of living matter, the application of this science in the end proved that, without a deepened cognition of the peculiarities of the state of living matter, the direct application of physico-chemical laws is also insufficient for the explanation of vital phenomena, that is, of the manifestations of living matter. Thus, at first the application of physical chemistry to vital phenomena led to the denial of the existence of a special molecular state in the molecules composing living matter, which was explained by the hope of managing here with the aid only of the physico-chemical laws of multiphase, highly disperse systems. But at the present time we are, it seems, living through a new turn and stand once again before a transformation, fraught with consequences, of our conceptions of the structure of living matter. This is called forth, on the one hand, by the development of experimental biology, which discovers ever new regularities and reveals ever more clearly the insufficiency of the application of physico-chemical laws without taking account of the special state of the molecules out of which living matter is built. On the other hand, the cause of this is the most recent achievements of physics, which give us methods for a deeper penetration into the intimate structure of matter and of molecules. If the last period of the application of physical and colloid chemistry proceeded, so to speak, under the sign of the liquid phase, while the solid phase—the protein micelles of colloids—was taken into account only in so far as, in consequence of the dissociation of its extreme acidic and basic groups, it carries on its surface a charge depending exclusively on the composition of the liquid phase, then it can hardly be doubted that, by means of the most recent conceptions and methods, the next period will approach the deepened investigation of precisely the solid phase and of the molecular forces acting within it, in order finally to find an explanation of vital phenomena in the special state of the molecular structure characteristic of living matter. The general laws of motion of living matter set forth by us require, as we showed at the beginning of this chapter, precisely such a development.
Let us now see what facts we already have at our disposal and what conception of the structure of living matter, answering to our general laws and permitting the explanation of vital phenomena, we are already able to form for ourselves.
We have already said that the non-equilibrium state must be given by the molecular structure itself. What facts speaking in favour of this do we already have at our disposal?
The most direct method that modern physics gives us for the study of the structure of matter is, as is known, the X-ray photograph, as well as investigation in polarised light and the study of absorption and emission spectra. Let us consider for the time being the still not numerous results obtained by these methods in application to living matter. The method of X-ray photographs, as is known, was applied to the study of living tissues by K. Meyer, who showed that the micelles forming the muscle fibril, or their basal corpuscles, possess an elongated form and curl up upon the contraction of the muscle. We shall consider these investigations and their significance for the mechanism of muscular contractions in more detail in the chapter on irritability. Here it is important for us that Meyer and Mark give the following conclusion in their book:[Author’s note:] K. Meyer and H. Mark, Der Aufbau der hochpolymeren organischen Naturstoffe, Leipzig, Akad. Verlag, p. 234, 1930. “On the basis of the present level of our knowledge we can say approximately the following: crystalline micelles, that is, micelles built in the form of a lattice, occur not in really living, but only in dead cells and their parts; they acquire a fully crystalline structure when the cells die.”
It is known that a lattice structure corresponds to the equilibrium state, that is, to the minimum of free energy. Thus the conclusion of the cited authors means only that, so long as the cells live, the structure of living matter is really not in equilibrium and that its state is conditioned by the molecular structure itself. The fact that upon the dying of cells a lattice structure arises and the micelles acquire full crystallinity speaks in favour of the view that the not-fully-crystalline structure corresponds not to a labile or metastable equilibrium, but precisely to a non-equilibrium state, that is, to such a state as is not viable without constant processes supplying energy, and without constant maintenance or renewal, as is also the case with respect to excited atoms and molecules. This result is fully confirmed by investigations of living tissues in polarised light. Anisotropy, double refraction, is exhibited by many tissues and parts of cells. But the double refraction which we observe in the parts of living cells is always a double refraction of form, as, for example, in the fibres of striated muscle or in other fibrous and elongated structural elements. It is conditioned not by an anisotropic, lattice structure of the separate micelles or molecules composing the given body, but is called forth by their elongated form and uniform orientation in the given body. Intrinsic double refraction, conditioned by the lattice structure of the micelles forming the tissue, occurs only in dying or already dead parts of the organism, as, for example, in the cellulose walls of plant cells, the lipoid granules of nerve sheaths, and so forth. The heads of spermatozoa, which according to Schmidt exhibit strong double refraction, are likewise no exception to this rule. This is evident from the fact that killing, boiling, fixation do not alter the degree and character of the double refraction of the heads. Schmidt ascribes this double refraction to nucleoproteids. But the investigations of Shinke and Shigenaga[Author’s note:] Shinke and Shigenaga, Cytologia, vol. 4, No. 2, 1933. showed that at least the chromosomes of plant cells contain nucleoproteids only in the spiral part, while the main mass contains not nucleoproteids but chiefly other proteins and lipoids. Extremely interesting is the following observation made in our laboratory. Spirostomum ambiguum usually exhibits no double refraction in the polarising microscope. If one cuts off a small piece from it, then the piece without a nucleus dies, while the one containing the nucleus continues to live and move. Sometimes one succeeds in observing how the whole dying portion suddenly becomes sharply doubly refracting, while the surviving piece remains optically isotropic. We see that here too, upon dying, an ordered structure appears, corresponding to the equilibrium state and calling forth anisotropy. It should be noted that the double refraction, upon dying, disappears again after a certain time; this is probably explained by the autolysis that sets in. If this supposition is borne out, then we obtain a fine optical method for determining the moment of the onset of autolysis—a method considerably surpassing the chemical methods in fineness.
But if the molecules and micelles of living matter do not possess a lattice structure, which appears only upon dying, then it follows from this that the micelle of living tissue must possess an electrical charge with respect to the micelle of non-living tissue. This is necessary because in a lattice structure the electrostatic forces saturate one another to the maximum. The groups of atoms or the ions of the lattice are, in accordance with the equilibrium state, at a minimal distance from one another. Every curving of such a lattice structure, accompanied by an increase of this distance and consequently by an increase of free energy, must also be accompanied by an increase of the strength of the electric field; or, in other words, the latter must diminish upon the onset of equilibrium, that is, upon the arising of the lattice structure, as is the case upon dying.
There is known a very widespread general phenomenon, according to which every living tissue, upon dying brought about by some local injury, receives a negative charge with respect to the uninjured, non-dying tissue: in other words, the potential falls from the living tissue to the dead. This rule holds for all living tissues, both plant and animal. The currents thus obtained are known under the name of “currents of rest” or “currents of injury.”
This phenomenon can be directly explained by means of our conception of the non-equilibrium state of living matter, conditioned by the non-equilibrium of the molecular structure itself, by the “curving” of its lattice constitution, and confirmed, as we see, by physical methods of investigation. The electrical difference of potential between living and dead tissue is thus the result of the difference of the molecular structures themselves.
It is known that at the present time physiology gives another explanation for these currents of rest. At the beginning of this chapter we pointed out that the modern physico-chemical explanation of the properties of living tissue does not recognise a special state of living matter and therefore reduces all phenomena of difference of potential to the properties of the boundaries of phases or of membranes, that is, to the boundary conditions; in accordance with this scheme, the general property of all living tissues of producing currents of rest is likewise explained. One theory proceeds from the existence of membranes [Bernstein] with different permeability for cations and anions; another [Beutner] from the properties of the lipoid sheath present on each cell facet as a special phase. But in order to explain why injured tissue is always charged negatively, one would of course have to suppose that all cells are always permeable predominantly to cations, and such a supposition seems to us nonetheless highly doubtful. We know cells, as for example the red blood corpuscles, with respect to which precisely the reverse is correct. As regards Beutner’s theory, it too is contradicted, again from the standpoint of modern theories of permeability, by the fact that the lipoids on the facets of cells do not form a continuous, unbroken phase. That the “curving” of the lattice structure does indeed lead to the arising of a positiveThe source reads «положительного» (“positive”), which is correct in context: the deformed (living-analogue) point is positive relative to the undeformed (lattice, dead-analogue) point, as the following experiments confirm. charge with respect to the non-curved part, of this we were able to convince ourselves in a series of experiments carried out in our laboratory jointly with V. A. Muzheev.
If one takes a copper or zinc wire and applies to two of its points non-polarising electrodes, as is done in physiology for leading off bioelectric currents, then a mirror galvanometer (of sensitivity 10⁻¹⁰) connected between the two points usually gives no deflection at all. If, however, one deforms some place on the wire, for example by twisting or flattening, and applies to it one of the electrodes, leaving the other at the undeformed point, then the galvanometer reveals a strong deflection. The magnitude of this deflection for copper wire many times exceeds the current of rest in the m. gastrocnemius of the frog, the deformed point being positive with respect to the undeformed. Thus the deformed place corresponds to the uninjured, living tissue, and the undeformed, that is, the place with a lattice structure, to the injured, dead tissue. This fully answers to our supposition that living molecules, or micelles, possess a deformed, non-equilibrium lattice structure, which also explains their positive charge with respect to dead tissue. That the positive charge of the deformed place is indeed called forth by the non-equilibrium state of the structure conditioned by the deformation, and does not correspond to a new equilibrium, is evident from the fact that the difference of potential equalises rather quickly and the galvanometer returns to the zero point. The course of this process of equalisation seems to us very instructive. If we are dealing with an ordinary copper wire, consisting of separate crystals, then in the majority of cases already after 5 minutes the galvanometer returns to the zero point, the fall proceeding not smoothly but accompanied by jumps in various directions. It is evident that we are here registering the restoration of the correct arrangement of the separate crystals with respect to one another. This is proved by an experiment carried out by us with a single crystal of cadmium. Here the deformed (flattened) place was likewise charged positively, but the difference of potential was preserved, with slight fluctuations, in the course of half an hour.
But not only the deformation of simple metallic crystals leads to a positive charge with respect to the undeformed place. The same thing we succeeded in observing also on protein gels. For this purpose we took a solution of gelatine in Ringer’s fluid, poured it into a Petri dish, and when it had solidified sufficiently and acquired the elasticity needed for the experiment, we cut strips out of it. If one applies electrodes to two points of such a gelatine strip, the galvanometer reveals no deflections; if, however, one of the points was subjected to stretching, then an insignificant but distinct deflection was observed, the stretched point again turning out to be positive, that is, corresponding to living, uninjured tissue, while the unstretched point corresponded to dying, injured tissue.
We see that the curvings, the deformations of the lattice structure brought about by external influences, both in metals and in protein bodies, lead to the appearance of a positive charge at the deformed place. This deformed state of the molecular or lattice structure is a non-equilibrium state, which equalises after the cessation of the action of the external, deforming forces. In living tissue, on the contrary, this non-equilibrium state, this deformed molecular structure, is stable in the sense of our general law of motion of living matter, that is, it is maintained or constantly restored at the cost of the energy of the continuous processes of equalisation proceeding in the living tissue. Therefore in living tissue, in which the processes of metabolism, that is, of the restoration or maintenance of this deformed state of the molecular structure of living matter, are disturbed or do not take place at all, an equilibrium lattice structure arises. Let us here briefly indicate that from our definition of stimulation, precisely as that which calls forth a disturbance of this maintenance of the non-equilibrium structure, it directly follows that at the stimulated place the deformation must diminish and the structure approach the equilibrium state. The stimulated place must, consequently, possess a negative charge with respect to the non-stimulated. That this is indeed so is sufficiently known from the physiology of stimulation. As regards the connection between the intensity of metabolism and the strength of the current of rest, it, as is evident from all that has been said above, must undoubtedly exist, and we shall return to it in more detail in the chapter on adaptation and irritability.
According to what has been said, the electrical difference of potential, or charge, on the various parts of living tissue must depend on the degree to which the molecules of the living substance are deformed, that is, on how much greater, in consequence of their excited non-equilibrium state, their free energy is in comparison with the unexcited state. That part of the free energy of unit mass which living substance possesses in consequence of its deformed, non-equilibrium state, we, by analogy with the chemical potential, call the potential of living substance and denote by µ.
The electrical potential must be the greater, the greater the distance from the lattice structure, that is, the stronger the deformations. However, in the chapter on metabolism it will be shown that, on the one hand, µ must diminish with growth, and on the other, that growth takes place where µ is great. There it is deduced that µ diminishes in the course of individual development and that its maximum lies at the beginning of this development, owing to which the growth of a newly arisen individual, of the sexual and embryonic cells, also becomes possible. But then, by virtue of all that has been said here, the degree of deformation of the lattice structure of the molecules of living tissue, and consequently the electrical charge as well, must here be strongest. That this is indeed so is shown by the measurements made by Lund on Obelia. His measurements show that in the formation of buds this organism exhibits a clear polar tendency. “At the same time an electrical polarity is observed in this organism: the growing apex is charged positively with respect to the other parts of the body.” Lund (1930) also adduces an example in which the vegetation points of Pseudotsuga mucronata are always positive with respect to the other parts of the body, if the electrodes are applied to the bark. “Within the tree, however, there prevails a fall of potential in the reverse direction: the base is charged positively with respect to the apex” (cited after Went, Jahrb. f. wiss. Botanik, Bd. 76, 1932).
Thus we see that the theory of living matter developed here fully corresponds to the conclusions following from our general laws of motion, and at the same time directly explains the regularities of bioelectric phenomena: when assimilation outweighs dissimilation by virtue of the higher potential of living matter, as is the case in embryonic tissues, buds, vegetation points, then we have before us a great deformation of the lattice structure, a greater distance from the equilibrium state, and by virtue of this such a place receives a positive charge. If, however, the processes of assimilation, that is, the maintenance or constant renewal of this non-equilibrium structure, are disturbed by stimulation, and if the structural energy diminishes, thereby approaching equilibrium, then we have in such a place a negative wave. Finally, if the tissue dies, an equilibrium lattice structure arises, and then this place is charged negatively with respect to the living parts of the tissue.
We have already said that the lattice structure of the molecules is not preserved after death, since the double refraction manifested upon dying gradually disappears again. We then already expressed the opinion that this is the result of chemical changes setting in in consequence of autolysis. To this fully corresponds the fact that the maximal negative charge of dead tissues is not preserved, but gradually becomes more positive,[Author’s note:] Cf. Bose, Électrophysiologie comparée, Gauthier-Villars et C-ie, p. 151, Paris, 1927. probably chiefly at the cost of the acidic products of autolysis. The appearance of intrinsic double refraction and of negative charge are phenomena accompanying the transition of the non-equilibrium structure of the molecules, characteristic of the living state, into the equilibrium lattice structure. The subsequent diminution of the double refraction and of the negative charge is the expression of the post-mortem but regularly setting-in processes of autolysis, of which we shall speak further on. If the currents of rest are explained not by a lipoid phase, in which various ions are differently soluble, and not by sheaths acting as a sieve for ions, but are based on a special state of the molecules of living tissue, then we must be in a position to demonstrate the presence of these currents even after the disturbance of membranes or of the continuity of the lipoid surface, and also in those cases where there can be no question of any membranes whatever. That this is indeed the case we succeeded in showing in our laboratory jointly with V. A. Muzheev. We set up our experiments on a pulp of muscle and on blood serum. In a frog both mm. gastrocnemii were dissected out, after which one of them was immediately killed by rapid immersion in hot Ringer’s solution. Then both muscles were ground in a mortar until they were completely converted into a pulp. With this pulp two small glass dishes were filled. Both pulps were joined by a glass tube filled with agar dissolved in Ringer’s solution and set, in order to close the circuit. Then into each of the pulps a non-polarisable electrode was immersed, which were connected with sensitive (10⁻¹¹) galvanometers. It thereby turned out that the pulp from living muscle always possessed a positive charge with respect to the pulp from dead muscle. In the same way it was possible to show that the pulp from a muscle fatigued by prolonged tetanic stimulation is always negative with respect to the pulp from an unfatigued symmetrical muscle. In view of the fact that the active reaction, especially in fatigue, shifts toward acidity, by which a current in the reverse direction ought to have been called forth; further, that we here have no membranes or cell boundaries, while the direction of the current is always the same as in the currents of rest—our experiments prove that the currents of rest are based not on an uneven distribution of ions by means of membranes and not on the action of a continuous lipoid phase, but on the molecular structure of living matter itself. It is quite understandable that the strength of the current thus obtained must be lower than in an undamaged muscle. In the first place, here, in view of the destruction of the cell structure, those energetic processes which maintain the non-equilibrium structure of the molecules are also disturbed, so that we have to do with an already dying tissue; in the second place, the charge or electrical polarity is strengthened owing to the fact that the molecules or micelles are arranged in the muscle fibre in the form of a chain (which also explains the double refraction of the fibre), by virtue of which the undisturbed structure directly leads to a strengthening of the current of rest.
It is interesting to note that this “current of rest” is observed only on the pulp of muscles of winter frogs, while in summer ones it could not be detected. This fully corresponds to those well-known profound changes in the physiological properties of the muscles of frogs which take place in connection with the time of year and hamper the investigation of the processes of stimulation in summer frogs.
In the same way we set up experiments on blood serum; each serum was divided into two portions, of which one was inactivated by heating at 56° for half an hour. The active serum was poured into one dish, the inactive into another; then both dishes were joined by a glass tube filled with agar, and the currents were led off to the galvanometer. In the majority of cases it again turned out that the active serum is charged positively with respect to the inactive. In individual cases no difference of potential was observed; but a reverse charging was never observed. As was to be expected, the deflections of the galvanometer here are still weaker. That we here too have to do not with the action of electrolytes and especially with the action of a changed concentration of hydrogen ions, is proved, in the first place, by precisely these weak deflections, and in the second place, by the fact that the arising difference of potential exhibits a far-reaching parallelism with the diminution of the surface tension of the serum upon its inactivation (see below).
From what has been said it follows that our conception, according to which the molecular structure of living matter is itself a non-equilibrium “curved” lattice, directly explains the electrical properties of living tissue, especially the appearance and direction of the currents of rest.
From this conception of ours concerning the state of the molecules forming living matter, there follows yet a whole series of consequences which we can likewise verify by experiment. Thus, for example, it is evident that if the lattice of the living molecules is deformed, and consequently the charges of the separate atoms or atomic groups are not at that minimal distance from one another which corresponds to equilibrium, then in such a case the dipole moment of the molecules must increase, if they possess one, or they acquire such a moment, if it is absent in the equilibrium state. Consequently, upon dying, that is, upon the transition of living matter into dead, in other words, in the equilibrium state, the dipole moment of the molecules must diminish. (This conclusion would be incorrect only in the case where we had to do with molecules in which atomic groups with equal positive and negative charges follow one another in alternation, and which by virtue of this in general possess no dipole moment, or else in the case where the deformation consisted in a uniform stretching of the whole molecule. But both these cases appear to be excluded.) Thus, living matter is electrically polarised, even without the application to it of an electric field from without, and the degree of this polarisation diminishes upon dying, that is, upon the onset of the equilibrium structure. Correspondingly, the dielectric constant must also diminish upon dying. Unfortunately, systematic measurements of the dielectric constant in living and dead tissue are still lacking. Only in Fürth[Author’s note:] Fürth, Annalen der Physik, IV Folge, Bd. 70, 1923. do we find some comparative measurements in active and inactive serum. In favour of our being entitled to regard blood serum, if not as living, then at least as surviving tissue, speaks a whole series of serious considerations which we shall yet touch upon further on. In any case, we know that the state in which the molecules of active serum are found is not an equilibrium one, since it is known that inactivation sets in even upon quiet standing at room temperature without any shaking or heating. Only in this case this process of equalisation lasts somewhat longer (2–3 days). Fürth’s measurements indeed show a fall of the dielectric constant of serum upon its inactivation. We illustrate this by two experiments from Fürth’s cited work (p. 55):
Experiment No. 40. Inactivated sera (man). One part of the serum remained unchanged (a), another was heated for half an hour to 50° (b), a third was subjected for half an hour to shaking (c): a .... E = 85.5 ± 1.0 b .... E = 83.3 ± 1.0 c .... E = 81.9 ± 1.0
Inactivated sera of the guinea-pig. The serum was treated in the same way as in Experiment No. 40.
a .... E = 85.2 ± 0.5 b .... E = 82.8 ± 0.5 c .... E = 82.2 ± 0.5
These experiments show that the dipole moment of the molecules found in serum really is lowered upon inactivation. That this lowering of the dipole moment is conditioned by a change of the molecular structure is especially distinctly evident from the fact that “the degree of dispersity, to a first approximation, apparently does not influence the dielectric constant.”[Author’s note:] E. A. Hafner, Ergebnisse der Physiologie, Bd. 24, p. 585, 1925. That we here have to do with a transition into the equilibrium state is evident from the fact that this process proceeds spontaneously without any influence from without.
We see that, in so far as measurements already exist, they confirm the conclusion from our theory of the non-equilibrium structure of the molecules of living matter concerning the lowering of the dipole moment, or electrical polarisation, of these molecules. A further conclusion can be drawn concerning the form of the molecules of living matter. Since equilibrium, generally speaking, is present at the minimal distance of oppositely charged atoms and atomic groups from one another, that is, at their closest arrangement, when the electrical forces are saturated to the maximum, while the dipole moment of one and the same atomic groups with the same charge depends to a great degree on the distance of the “centre of gravity” of the negative and positive charges, then it is evident that in general we shall be the further from equilibrium, the greater this distance. We must consequently assume that the chains forming the living molecule are stretched out in length and that upon dying they curl up and shorten. The molecules of living matter must, consequently, possess an elongated form, this elongated form being conditioned by the non-equilibrium of their structure and, upon dying, that is, upon the transition to equilibrium, changing in the sense of shortening.
There are not a few data speaking for the view that the molecular building-blocks out of which living matter is built possess an elongated form. Let us point to the work of Seifriz,[Author’s note:] Seifriz, The American Naturalist, v. 63, No. 688, 1929. in which he arrives at this conclusion on the basis of a series of physical properties of living matter. Seifriz writes: “The quality of contractility of such gels as cellulose, rubber, gelatine, and protoplasm is present everywhere where there are elongated and stretched structural elements.” But for us it is here important that this elongated form of the molecules of living substance corresponds precisely to the non-equilibrium structure of them and must be maintained by the processes of metabolism supplying energy, in consequence of which it must, upon dying, change in the sense of shortening. For this we also have at our disposal certain observations directly proving our point of view. Let us adduce here, first of all, once again the X-ray photographs by K. Meyer of a resting and a contracted muscle. Upon stimulation and dying the muscle contracts, and this process is conditioned precisely by the contraction of the micelles forming the muscle fibre. Muscle tissue, as it were, makes use of this property of the molecular structure of living matter in order, upon the disturbance of it, to perform external work in the sense of our principle of the work of the structural forces. But the investigations of Edsall and Muralt[Author’s note:] Edsall and Muralt, The Journal of Biological Chemistry, Vol. 89, 1930. of solutions of myosin also confirm our point of view. These authors found that, upon careful and gentle preparation of solutions of myosin, the latter exhibit “double refraction of flow,” and this proves that under these conditions the elongated form of the micelles is preserved. Further, it turned out that this double refraction of flow disappears already upon the slightest changes of reaction and various other influences. For us especially interesting and important is the
observation that even if one simply lets the solution stand, the double refraction disappears, and indeed no other signs of denaturation appear.
This shows once again that the elongated form corresponds to the non-equilibrium state and that the first change setting in spontaneously, in the case where the constant processes supplying energy for the maintenance or restoration of this state are lacking, is accompanied by a shortening of the micelles. This elongated form of the molecules or micelles of living matter is of great significance, if one takes into account that dipoles can associate in the form of chains, in consequence of which there arise formations with large dipole moments, with strong electrical polarity. The microscopic structure of many cellular formations of the kind of mitochondria and contractile elements, of muscle fibres, is very probably the result of such an association of the molecules of living matter. In view of the fact that these associations are conditioned by the large dipole moment and elongated form, which in turn are based on the non-equilibrium state of the molecules, it becomes clear to us in this—and only in this—way why, upon the dying of the cell, these smallest elements of microscopic structure also disappear: the mitochondria disappear, the fibrils of muscle and nerve fibres yield place to a granular structure, and so forth. (To avoid misunderstandings let us emphasise once more that we are here speaking of dying, and not of killing [умерщвление — umershchvlenie] by fixation. In the first case there takes place a process of equalisation with transition into the equilibrium state; in the second, however—and in this consists, for the histologist, the art of choosing suitable fixatives—the whole question consists in achieving such changes of conditions under which these non-equilibrium structures are fixed in an equilibrium position. In our opinion the theory of fixatives ought likewise to be based on the conceptions and facts set forth here, or developed in this direction.)
In the fluids of the body, where there is no structure in the histological sense, but where we must assume the presence of molecules of living substance still in a non-equilibrium state, as, for example, in blood serum, we can expect an ordered distribution of the molecules only in the boundary layer. In it the distribution or association of the molecules will be according to the following scheme:
Consequently, the electrostatic attraction, or the density of distribution of the molecules in the boundary layer, will be the greater, the stronger the dipole moment of the molecules; if the latter diminishes, then the striving toward association diminishes, and consequently the surface tension as well. Indeed we know that upon the inactivation of serum, simultaneously with the mentioned lowering of the dielectric constant, the surface tension also diminishes, and indeed this takes place even at room temperature without any external influence. Consequently, such a long-known phenomenon as the lowering of surface tension upon the inactivation of blood serum likewise receives a direct explanation by means of our conceptions of the structure of living matter. This is the more interesting in that no other explanation of this phenomenon exists, although it has been subjected to detailed study, especially by du Noüy. Indeed, standing or warming, leading to inactivation, call forth at the same time a diminution of the degree of dispersity of the protein bodies of the serum, and this ought to have led to a raising of the surface tension. Here, it is true, one might suppose that upon inactivation surface-active substances adsorbed on the protein micelle are liberated. But against this speaks a number of facts. It is known that by means of freezing with subsequent thawing one can liberate substances adsorbed on the surface of colloids. G. Grinberg, in our laboratory, at the suggestion of I. Weichherz, carried out corresponding experiments on blood serum. The results he obtained show that the diminution of the surface tension of serum upon inactivation cannot be explained by the formation of surface-active substances.
In Grinberg’s work some other phenomena are adduced as well, likewise contradicting this supposition. Let us here indicate yet one more regularity, which, on the one hand, makes this supposition very little probable, and, on the other, can be satisfactorily explained only by means of the conceptions developed by us here. To this regularity we have already pointed in one of our earlier works.[Author’s note:] E. S. Bauer, Biochem. Zeitschrift, Bd. 138, 4/6, 1923. This regularity consists in the fact that the differences in surface tension in various active sera, based on differences in age, constitution, and so forth, disappear after inactivation, that is, the equilibrium state toward which the various sera strive is the same for all of them.
In our opinion this regularity speaks decisively in favour of the conceptions set forth here concerning the non-equilibrium structure of the molecules of living matter, and against the arising or liberation of any surface-active substances whatever. Were the latter the case, then of course the magnitudes would have to exhibit large fluctuations. If, however, we have to do with a state, deviating from equilibrium, of one and the same kind of molecule, then, depending on the magnitude of the deviation from the equilibrium state, large fluctuations must be observed, disappearing after the onset of equilibrium.
Another, very important observation was recently made by Vlès and Gex,[Author’s note:] Vlès et Gex, Archives de Physique biologique, vol. 6, p. 255, 1930 (cited after Needham, Chemical Embryology, vol. I, 335, 1931). which can likewise be regarded as a direct proof of the view that the non-equilibrium of living matter is conditioned by the molecular structure itself and that the latter changes upon the onset of equilibrium, that is, upon dying. These authors investigated the absorption spectrum of normal sea-urchin eggs and found that in them the absorption bands characteristic of proteins in the ultraviolet, especially between 2600 and 2400 Å, are absent. Upon the onset of cytolysis, however, that is, upon dying, the absorption spectrum typical of proteins appeared. The supposition that the absorption spectrum of proteins was in some way masked in the living egg seems to us entirely unjustified and unfounded, all the more since we cannot adduce any analogies for such a phenomenon but, on the contrary, know that no one substance ever disturbs the spectrum of another, and that the whole of spectral analysis is based precisely on this possibility of the simultaneous obtaining of the spectra of all the components of a given substance. The result obtained by Vlès and Gex is a direct proof of the view that the molecules out of which protein or living matter is built possess a different structure than in the non-living state, and that upon dying this structure is transformed into the one we know in protein molecules outside the organism. We know that absorption bands, in contrast to lines, are conditioned by molecules. On the other hand, it is clear that if a molecule in the living state does not absorb, but in the dead, on the contrary, does absorb some wavelength, then upon the transition from the first state into the second it must emit precisely this wavelength. This gives us the possibility of calculating, by means of Bohr’s fundamental equation, the energy that the molecule of living matter gives off upon the transition into the equilibrium state, that is, upon dying.
On the basis of what has been said we write:
where denotes the energy of the molecules in the living state, and in the dead, that is, in equilibrium, is Planck’s quantum of action, and the frequency of the oscillations of the emitted rays. On the basis of the experiments of Vlès and Gex we substitute for the magnitude corresponding to a wavelength of 2500 Å, that is,
and since erg/sec., we calculate from this:
for one molecule upon the transition from the living state into the dead. Taking now the molecular weight as equal to 20000 (the molecular weight of haemoglobin is about 16000), we obtain for the mass of one molecule:
and the number of molecules per 1 g. of protein:
hence the energy upon the transition of 1 g. of protein from the living state into the dead equals:
Had we taken the molecular weight as equal not to 20000 but to 10000, then we should have obtained not 5 g-cal. but 10; at a molecular weight of 15000 we should have obtained 7 g-cal. Had we taken a wavelength of 2400 Å instead of 2500, then the order of magnitude would likewise not have changed. Thus, this calculation leads us to the conclusion that upon the transition of 1 g. of living substance into the state of equilibrium, that is, upon its dying, a quantity of energy of the order of 10 g-cal. must be liberated. As regards the experimental data on the evolution of heat upon dying, on this question there are as yet no investigations on the basis of which the correctness of our calculation could be verified. The very few measurements made by Meyerhof, Lepeshkin, and others gave contradictory results. Meyerhof found no evolution of heat in well-respiring red blood corpuscles after killing them with acrolein. Lepeshkin, killing yeast cells with corrosive sublimate and chloroform, found an evolution of heat which on the average exceeded the evolution obtained upon the precipitation of the protein of yeast by the same means by 2 g-cal. per 1 g. of protein. Lepeshkin asserts that he found the same thermal effect in red corpuscles as well. But these experiments are unsuitable for the resolution of the question of the existence and magnitude of the difference in energy of living and dead matter. Experiments with the killing of cells or tissues are essentially incorrect and in general cannot give an answer to our question. We have already, in connection with the question of the change of the microscopic structure of tissues, mentioned that in the thermodynamic respect too there is an essential difference between dying and killing. In the first case we have a process of equalisation, in which the system, being in non-equilibrium, passes over into the state of equilibrium determined by the given conditions of the system. In the second, however, we essentially and, moreover, in a rather indeterminate manner change the conditions of the system and establish some equilibrium state. It is clear that any at all unambiguous answer to our question we shall obtain only by measuring the evolution of heat upon dying. If, however, we set about killing the tissue, then we could expect a positive effect only in the case where we proceeded from the previously set forth old conception, according to which in living tissue there is some labile substance which decomposes upon every mode of death and always passes over into one and the same, less energy-rich compounds. We have already said that this conception is incorrect and is at present rejected. If, however, one holds that the conditions of the system, or the structural forces, perform no work, since the constituent parts of the system themselves, being in equilibrium and therefore fulfilling only the role of transformers, do not undergo changes of their state, then of course it is as hard to expect an evolution of heat in consequence of killing as in consequence of the stopping of a machine, and we must come to the denial of structural energy, as Meyerhof indeed does. But here it is necessary to remember only that experiments with killing in no measure speak for or against the existence of structural energy. A positive result upon dying could, from this point of view, be explained as a temporary “idle run” at the cost of the processes supplying energy. In other words, the evolution of heat could be interpreted in such a way that it is conditioned by enzymatic cleavage, for example of glucose, by oxidation, and so forth, taking place, however, without the slightest changes of structure under the given conditions. The process of dying would then be nothing other than the conclusion of the chemical processes in a system where equilibrium has not yet set in, but where the conditions of the system, that is, the structures, do not undergo change. We have already sufficiently shown that this is incorrect and contradicts all the facts. If we wish to measure the energy liberated in consequence of the change of state of the structures upon their transition to equilibrium, then, in the first place, we must measure it upon dying, and not upon killing, and, in the second place, the method of the calorimeter is for this unsuitable, since we here have to do not with a rapid chemical reaction, similar to an explosion, but with a process of equalisation which can last for hours, in view of the fact that, as we know, the processes proceeding in living tissue and supplying energy retard precisely this equalisation. Finally, it still remains to be shown that the heat given off is greater than would correspond to the processes of cleavage proceeding in the tissue and likewise supplying energy. This excess will then be the measure of the free energy which falls to the share of the matter in the living state and can diminish without change of conditions, that is, in consequence of the non-equilibrium nature of this state.
Such measurements of the heat formed upon dying, satisfying the indicated requirements, were indeed made by Hill.[Author’s note:] A. V. Hill, Proc. of the Royal Society, B, vol. 103, 1928. In connection with the study of heat formation at rest and its dependence on the consumption of oxygen, Hill found that the quantity of heat given off by a resting muscle in the absence of oxygen over 24 hours is so great that it can in no way be explained by means of the normal processes supplying energy, such as glycolysis, the breakdown of creatine-phosphoric acid, and so forth. Moreover, the formation of heat continues for several hours after the muscle has lost its irritability, and even rises somewhat toward the end. Consequently, we here really have to do with heat formation in living tissue upon its dying, and find that the quantity of heat liberated upon dying many times exceeds that which the energetic processes known to us, proceeding during dying, yield. According to Hill, over 8 hours 5 g-cal. per 1 g. of muscle are formed. What is the nature of those processes which yield such quantities of energy upon dying remains unclarified. Hill gives the following explanation of his data: oxidation is necessary for the preservation of the normal cell structure. At a definite degree of oxygen deficiency a disturbance of the structure sets in, leading to the result that substances incapable of reacting with one another in the normal structure now begin to react freely, “and the organised system is rapidly transformed into a chaos of biochemical processes.”[Author’s note:] Loc. cit. The structures which prevent these chaotic biochemical processes and which must be preserved are membranes and boundary layers. “Thus we must strive to seek the retarding agents, which maintain the dynamic equilibrium of life and do not permit the realisation of undesirable reactions, not in some mysterious chemical control taking place in a homogeneous medium, but in really existing, material membranes and boundary layers.”[Author’s note:] Ibid. Of course it is not by chance that here there arises once again the same question: boundary layers and membranes separating various phases from one another, or processes within one, homogeneous phase. In accordance with the conception prevailing at present, Hill denies the latter possibility. We have already shown that if we accept the non-equilibrium state of the system and transfer the immediate source of work for the preservation of this non-equilibrium into the system itself—and this is precisely what is characteristic of life—then we are compelled to ascribe the non-equilibrium state to the molecular structure itself. We here have to do not with a dynamic equilibrium, the presence of which Hill supposes, and this is best shown by his own experiments. Indeed, a dynamic equilibrium either requires no processes at all supplying energy for its preservation, or the source of them lies outside the system. But Hill himself is compelled to accept the existence of a non-equilibrium structure maintained by oxidative processes supplying energy; he himself speaks of the “normal microphysical architecture of the cell.” Upon dying this structure disappears; consequently, a certain quantity of heat must be liberated at the cost of the free energy of this state of microphysical architecture. Of course, it is not excluded and is even probable that such a change of structure will simultaneously call forth a shift of chemical equilibrium as well, since the latter depends, after all, on the conditions of the system. We shall occupy ourselves with this question further on in connection with autolysis. Here, however, we shall establish only the following: Hill’s experiments prove that upon dying an evolution of heat takes place, that the quantity of this heat considerably exceeds that which the processes of cleavage proceeding upon dying produce, and that part of this excess, or the whole of this excess, must be referred to the account of the changes of state of living matter taking place upon equalisation. The difficulty in the quantitative measurement of the structural energy liberated upon the transition
from the living state into the dead consists, consequently, not in the impossibility of demonstrating this evolution of heat but, on the contrary, in the fact that, in view of the large quantity of heat evolved, we are not in a position to decide whether we are entitled to refer all of it to the account of this energy of state, or whether we must ascribe at least part of it to processes of cleavage as yet unknown to us. If we apply Hill’s data in connection with our calculations adduced above for the resolution of this question, then we arrive at the following conclusion. A dying muscle gives off, per 1 g., 5 cal. over 8 hours; after approximately 16 hours it loses its irritability. Since the curve of heat evolution is almost rectilinear, we may assume that by this time 10 cal. have been evolved. To the account of the formation of lactic acid one can refer at most 2 cal., since 1 g. of lactic acid yields 385 cal., while at a 0.5% content of lactic acid, corresponding to 1.9 cal. per 1 g. of muscle substance, rigor of the muscle already sets in. If we refer a further 2 cal. to the account of the breakdown of creatine-phosphoric acid and the like, then we shall still be left with 6 cal. per 1 g. of muscle substance. But Hill’s data relate to the whole substance of the muscle, and, as is known, the muscle contains only about 17% dry substance; consequently, for living substance we must assume at most 15%, in order to be able to compare these measurements with our calculations. This amounts to 40 cal. per 1 g. of living substance, that is, eight times more than the quantity of energy calculated by us. From this it follows that either in the dying muscle, besides the processes known to us supplying energy, there proceed still others, hitherto unknown processes of cleavage, to the account of which we must refer this extraordinarily large quantity of energy, equal to 35 cal., or else the whole heat must be ascribed to that energy which is liberated upon the transition of the molecules from the living state into the dead. As regards the first possibility, not a single such reaction is known to us. Hill, however, accepts their existence; he even must accept it and must, by means of these unknown, hypothetical reactions, explain the whole excess of energy, since this is the inevitable consequence of the denial of a special non-equilibrium state in the molecular structure of living matter.[Author’s note:] Later Hill, it seems, was inclined to explain this heat formation by the equalisation of the vapour pressure of water in the chamber. Against this explanation speaks the course of the curve, which is almost linear over the span of 24 hours, and at the end even rises still further. Moreover, Hill makes this supposition only with respect to a muscle fatigued by stimulation. Therefore he also concludes his work with the proposal to seek these reactions. As regards the second possibility, namely that the whole quantity of heat should be referred to the account of the energy liberated upon the transition into the dead state, here the following must be said. We calculated the magnitude of 5 cal., proceeding from the supposition that the molecular weight of the molecules of which living matter consists is equal to 20000. This supposition, however, is to a certain degree arbitrary. It is based in turn on the supposition that protein molecules possess a molecular weight of this order of magnitude. But we know that determinations of the molecular weight of protein bodies give various results upon the application of various methods. Thus, determinations based on X-ray photographs give considerably smaller values of the molecular weight. We have grounds to suppose that the large molecular weight is obtained in consequence of various atomic groups adsorbed and bound by secondary valencies, and also by way of aggregation; proceeding from this, we should have had, in our calculations, to assume a considerably smaller molecular weight, which gives a greater evolution of heat upon dying, that is, upon the transition of the molecule from the non-equilibrium, deformed state into the equilibrium one. If we wished to explain the evolution of heat found by Hill of approximately 40 cal. exclusively by means of the difference of free energy of the molecules in the “deformed” and equilibrium states, then we should have had to assume a molecular weight equal to approximately 2500—a supposition which likewise cannot be considered excluded. Systematic investigations of various tissues in various physiological states and a deeper penetration into the structure of the molecules in the living state must resolve this question.
Important is the question of the presence and magnitude of that excess of energy by which the living state of the molecules is distinguished in comparison with the equilibrium, that is, dead, state—in other words, the question of the magnitude of the “structural energy.” This question is of essential significance in connection with our principle, according to which all external work is performed precisely at the cost of this structural energy, that is, by the systemic forces. This question is especially important also because, against the supposition of the existence of a special labile, energy-rich substance, the objection is constantly put forward that in such a case the evolution of heat upon dying would have to be very considerable. We have already said that this conception explains nothing, and in particular cannot explain the process of metabolism in the state of rest. It is quite unclear what processes proceed and what work is performed at the cost of the energy liberated upon the “explosion” of this labile substance. Therefore it also remains unclear what quantity of energy we ought properly to expect upon such an “explosion.” But if we proceed from our conception, according to which the food taken in—such as carbohydrates, proteins, fats—must first assume the state of the molecular structure of the living molecule, that is, be assimilated, and only by this roundabout path can liberate its energy; that is, that there exists only a plastic metabolism and all the energy liberated in this goes to the maintenance and preservation of this structure, while all the external work of the organism is performed at the cost of structural energy—if we proceed from this supposition, then we obtain an entirely different picture. There arises then the question: will 5 or even 40 cal. per 1 g. of living substance suffice to cover the external work performed by living systems? In comparison with the energy liberated upon the combustion of 1 g. of protein (over 4000 cal.), this energy can of course be neglected. But if we take some living tissue, for example the gastrocnemius of the frog, of a weight of 1 g. and with a protein content of only 10%, and assume for the structural energy only 5 cal. per 1 g. of protein, then this will correspond to work of 0.2 kilogram-metre (since 1 cal. corresponds to 0.427 kilogram-metre). At the cost of this structural energy, consequently, the muscle could lift a weight of 20 kg. by 1 cm. To the same results we come upon investigating the work of the heart in its contraction; for the human heart we obtain work of several hundred kilogram-metres, which can be performed at the cost of this structural energy. We see thus that not only does the whole structural energy calculated by us, proper to the “curved” non-equilibrium structure of living molecules, suffice to cover, in accordance with our principle, the whole external work of the organism, but that for this even an insignificant part of it is sufficient. This is understandable, since we have already said (and shall yet show in detail in the chapter on stimulation) that external work is made possible in consequence of a disturbance, a stimulation, coming from without, the structure of the molecules thereby approaching equilibrium, but the tissue by no means having to die completely, since then it would be impossible quickly to restore the capacity for work. The quantity of energy liberated upon oxidative breakdown, upon glycolysis, and upon other processes supplying energy, enormous in comparison with the structural energy and the external work performed by living tissues, is necessary for the maintenance, restoration, and new formation of the work-capable structure, that is, of living tissue. However, we shall further see that such processes supplying energy are possible only on the basis of this molecular structure in the living state.
But before showing how the theory of the state of living matter developed here permits us to explain the phenomena of metabolism, we should like to touch upon one conclusion from the conceptions set forth here. We have already pointed out that, on the basis of our conceptions and the above-mentioned experiments of Vlès and Gex, it must be accepted that upon the transition of the molecules of living matter from the deformed non-equilibrium into the equilibrium state, that is, upon dying, a radiation of a long wave, approximately from 2400 to 2600 Å, must take place. The existence of such ultraviolet rays of a somewhat smaller wavelength is indeed confirmed by Gurwitsch and his school under the name of “mitogenetic rays.” Concerning these rays there already exists an abundant literature. Very many investigators, however, relate extremely sceptically to the existence of “mitogenetic rays.” The experimental data of Gurwitsch himself contain within themselves not a few unresolved contradictions. The objective registration of the radiation by physical methods has hitherto not succeeded, and although in the latest experiments of Seyfert, as also of E. Lorenz,[Author’s note:] E. Lorenz, The Journal of General Physiology, vol. 17, No. 6, 1934. it was possible to register an intensity of 10 quanta per 1 second per 1 cm², yet all the experiments gave negative results. We shall not here discuss the whole literature and all the data for and against the existence of mitogenetic rays, but we should like to emphasise that, on the basis of the conceptions developed here, the existence of such a radiation can theoretically be expected precisely in this region of the spectrum. Even more important is the fact that a series of contradictions present in the data on “mitogenetic rays” can be resolved if, from the conceptions developed in the present chapter, we draw certain conclusions concerning the nature of this radiation. Above we said that the molecules of living matter possess, in consequence of their deformation, a greater dipole moment than would correspond to the equilibrium state, which also explains, for example, the high dielectric constant of serum and its lowering upon inactivation. We also pointed out that in the living state these dipoles are usually associated with one another. Thus, living matter is electrically polarised, as if it were under the influence of an external electrostatic field. From this it follows that such a non-equilibrium state of the structure of living matter conditions the presence of an electrostatic field in the latter. If, then, upon approaching equilibrium the molecules really emit rays, then this radiation takes place in an electrostatic field and must therefore be polarised. The possibility of a biological effect at so insignificant an intensity of radiation can be explained in this way, just as can the unreliability or impossibility of registration by means of the photoelectric effect. The fact that stimulation, that is, the disturbance of the molecular structure of living tissue, is achieved by means of such insignificant intensities, can be explained by the fact that polarised light calls forth a disturbance of the structure at considerably smaller intensities than non-polarised, since here the oscillations take place in one plane and consequently must shake the molecular structure more strongly. The supposition that polarised light can strongly influence certain biological processes already at minimal intensity has been expressed more than once. Some authors have attempted to connect various biological phenomena, the periodicity of which exhibits coincidence with the periodicity of the moon (for example, the reproduction of diatoms, the hydrolysis of starch in plants taking place at night, and the like), with the maximal polarisation of moonlight during these periods. Thus, Philip[Author’s note:] Philip, Nature, 130, 665, October 29, 1932. found that diatoms illuminated by day and kept in darkness at night reproduce considerably more slowly than those which were subjected at night to the action of moonlight; he considers it possible to explain this phenomenon by the strong polarisation of moonlight. Further, Semmens[Author’s note:] E. Sidney Semmens, Nature, 130, 243, August 13, 1932. showed that if one half of the leaves of Tropaeolum and other plants be covered with tinfoil, and the other subjected to the action of moonlight, then in the illuminated half the starch disappears sooner than in the unilluminated, if the experiment was carried out in the period of greatest polarisation of moonlight. On the contrary, no difference was observed if the experiment took place in the period of full moon, that is, at the minimal polarisation of moonlight. But all these experiments give, at best, only indirect data concerning the significance of polarised light for certain biological processes. An exact experimental investigation of this question does not yet exist; it will be carried out in the near future in our laboratory. But it should be noted that a stronger action of polarised light on the structures of living tissue, in comparison with non-polarised, is, it seems, observed for X-rays as well. In favour of this speaks clearly, in our opinion, the following observation made by Shimoda,[Author’s note:] Cited after Vendrovsky, Arch. f. exp. Zellforschung, Bd. XIV, 1933. who found that “tissue cultures of the heart of a large embryo are essentially damaged, and under determinable conditions even killed, by secondary X-rays. The primary rays, however, even in maximal doses produce no action on plants.” He thereby specially established that the damaging action belongs not to the secondary radiation of electrons, but to the secondary characteristic radiation.
But we know that the secondary characteristic radiation is fully polarised. The great difference in action found by Shimoda can hardly be explained by anything other than this circumstance.
If it is confirmed that polarised light exerts a special influence on living tissues, then the biological action of the ultraviolet rays issuing from tissues would also be explicable, in spite of their extremely insignificant intensity. As regards the unreliability or impossibility of registration by means of the photoelectric effect, this too would be understandable, since it is known that the measurement of polarised light by means of the photoelectric effect gives a result depending to a considerable degree on the chance direction of the ray. The probability that there will be obtained such a direction of the ray at which the plane of the electric vector will stand exactly perpendicular to the plane of the metal, that is, that the whole intensity will be registered, is evidently very small.
We see thus that the conceptions developed in the present chapter concerning the structure or state of living matter can, it seems, contribute to the resolution of certain contradictions that have arisen in the field of the study of “mitogenetic rays.” Theoretically, as we have shown, on the basis of the theory of living matter set forth, such a polarised radiation of precisely this wavelength is to be expected even independently of the experimental material concerning “mitogenetic rays.” According to our theory the radiation must set in in the case where the molecules of living matter pass over from their deformed, non-equilibrium state into the state of equilibrium, or approach it. However, this process must proceed constantly, since dissimilation begins with it; it is, so to speak, the very first phase of dissimilation. Some molecules bind by their basic or secondary valencies the compounds to be assimilated from the food; thereby they correspondingly rebuild themselves and approach the equilibrium state, while others are brought into a deformed state. Consequently, radiation is to be expected only when the processes of equalisation, of dissimilation, that is, of transition into the equilibrium state, begin to dominate, and assimilation is at least temporarily switched off, that is, when the structural energy proper to the deformed non-equilibrium state of the molecules is liberated throughout the whole tissue. This will take place upon dying or upon disturbance by stimulations, which retard the processes leading to the preservation of non-equilibrium and thus liberate the structural energy for external work; thus, for example, upon the contraction of a muscle these disturbances lead to an approach to the equilibrium structure of the molecules and are expressed, as we have seen, in the negative electrical charge of the corresponding parts of the tissue, for example of the nerve. To this, it seems, fully corresponds the fact that the so-called “mitogenetic rays” are observed precisely in such cases. There radiates blood, which, as we have shown, is inactivated spontaneously, with a lowering of the surface tension and of the dielectric constant, indicating that the deformed molecules with their large dipole moment pass over into the equilibrium state. Further, there radiate parts of tissues in which necrobiotic processes prevail, contracting muscles and nerves at the place of the “wave of excitation.” On the contrary, muscles and nerve tissue at rest do not radiate. Here it will be in place to point to yet the following circumstance. As we have shown, our theory requires that the radiation should take place upon the transition of living deformed molecules into the equilibrium state, should lie in the region of the ultraviolet part of the spectrum, and should be polarised in consequence of the electrical polarity of living structures. In living tissue this process is always bound up with the process of assimilation, that is, with the formation of new deformed molecules. We know that one of the most incomprehensible processes of metabolism in the living organism is the arising of optically active modifications of various compounds. It is known that synthetically one can obtain only racemic compounds; for the obtaining of optically active modifications one must have recourse to the aid either of already optically active compounds or of living beings and their ferments. But in recent times Kuhn succeeded in obtaining optically active compounds from racemic ones by means of elliptically polarised light. Therefore it is not excluded that in the arising of optically active modifications in metabolism it is precisely our radiation that plays a role.
Hitherto we have considered only the physical phenomena characterising the state of the molecules of living tissue in comparison with the equilibrium state after death (for example, X-ray photographs, properties in polarised light, electrical charge, dielectric constant, surface tension), or accompanying the process of equalisation upon the transition into the equilibrium state, such as heat formation, radiation, and the change of the above-named magnitudes. The question arises, can one now, on the basis of the conceptions developed here, draw any conclusions also concerning chemical changes, or, in other words, how can one explain by means of these conceptions those chemical processes which arise in the course of metabolism in living tissues, and how in general are we to conceive metabolism on their basis.
A molecule which is “demorphed,”“Demorphed” renders Bauer’s coinage «деморфирована» (from the Greek morphē, “form”) — a molecule whose form/structure has been disturbed; he uses it as a stylistic variant of “deformed.” that is, in which the electrostatic forces possessed by the separate atoms or groups of atoms (radicals) carrying a charge are arranged not in accordance with equilibrium, must, according to modern conceptions, possess such unsaturated, free, or secondary “residual” valencies as the molecule in its equilibrium state does not possess. The greater free energy of these molecules, arising in consequence of their deformed non-equilibrium structure, signifies, consequently, in the chemical respect a greater activity, a greater capacity of these molecules to react. In the language of theoretical chemistry this means that 1) these molecules, even before collision with the molecule entering into reaction, possess energy of activation, and 2) their constant of action is greater than in the state of equilibrium. By energy of activation is understood “that quantity of energy by which the formations situated in reaction, that is, in the state of maximal potential energy, exceed the average energy of the initial substances.”[Author’s note:] W. Hückel, Theoretische Grundlagen der organischen Chemie, Bd. II, p. 227 and following, Leipzig, Acad. Verlagsges., 1931. The constant of action, in turn, “indicates how many collisions, out of the number of those for which the energy of activation introduced by the molecules suffices, turn out to be successful. It stands in connection with the constitution of the molecule.”[Author’s note:] Ibid. It is quite evident that if in a reaction one of the participating molecules is supplied with a large quantity of unsaturated residual valencies, then the constant of action will be greater than if in the same reaction this molecule possessed a smaller number of unsaturated valencies. It is even possible that in the latter case the reaction in general cannot take place, while in the former it will proceed at an appreciable rate. This will always be the case when the corresponding molecules in their equilibrium state (as they exist outside living matter) need to have energy added in order that they may enter into reaction, while the energy arising from the collision of the molecule is for this insufficient. But if under the same conditions the molecule is deformed, then the “regions sensitive to impacts” are considerably larger in consequence of the free valencies, and the reaction will be able to set in. The same is the case with the energy of activation too. Since deformed molecules already possess, in consequence of their non-equilibrium structure, a greater potential energy, this precisely means that they, properly speaking, possess energy of activation even before collision; in other words, that the reaction tangle forming upon collision possesses a greater energy than the average energy of the initial and final substances in the equilibrium state, even independently of the energy introduced by the collision. Thus, in this case the reaction will take place even when, with undeformed molecules, it would have to be absent.
Thus, our theory of the state of the molecules of living matter gives us a direct explanation for the fact that almost all reactions taking place in the course of metabolism in living tissues do not take place outside the organism under the very same conditions of temperature, pressure, acidity, and so forth. Biochemists striving to explain the phenomena proceeding in the living organism without the admission of a special state of the molecules of living matter endeavour here too to find a way out of the difficulty by means of surface catalyses or the admission that there exist as many ferments as they find reactions. They are thereby still compelled to suppose the existence of a marvellous, harmonious joint action of the most various ferments, as is, for example, the case with respect to the alcoholic fermentation of zymase. All these admissions become superfluous if we proceed from our theory.
Let us note further that from our theory it follows that the deformed molecules of living tissue possess a greater dipole moment than would correspond to the equilibrium structure. But “an increase of the dipole moment can in itself increase the rate of reaction without dissociation of the substances participating in the reaction into ions. In principle one must admit the same possibility of the reaction of the dipole moment with another molecule for the halides and esters of other strong acids as well, and thus one can imagine that not all the reactions in which, in the presence of catalysts, substances react in the form of ions, will, in their absence, likewise proceed as ionic reactions.”[Author’s note:] Hückel, l. c., p. 268, as well as chapter 18, § 4, Dipole Moment and Rate of Reaction, p. 264 (literature there too).
We indicate this conclusion only briefly, in order to show that not only can the admission of a special deformed non-equilibrium state of the molecules of living matter be proved by a series of physical methods of which we spoke earlier, but that the chemical reactions and their course in the process of metabolism will probably also find their explanation on the basis of our theory.
The considerations set forth here on the kinetics of reactions give us the possibility of verifying, from this side as well, our calculations of the energy of living matter in comparison with dead, that is, of that energy which we have called “structural.” The energy of activation of some reaction is, after all, nothing other than that magnitude by which the energy of the reacting molecules exceeds the energy of the molecules in the inactive state. Consequently, a molecule of living matter capable of reacting and active must possess, in its deformed state, approximately such a quantity of “structural energy” as corresponds to the energy of activation of the reactions usually studied. The energy of activation is calculated by the formula:
where and denote the constants of the rates of reaction at temperature and , and the gas constant. For a temperature coefficient and for a temperature of 300° we obtain cal. per mole. If we again take the molecular weight of living matter as equal to 20000, then we obtain cal.; at a molecular weight equal to 2500, we obtain cal. We obtain the following result:
| Molecular weight | Structural energy, calculated by Bohr’s formula (in cal.) | Energy of activation (in cal.) |
|---|---|---|
| 20000 | 4 | 1.4 |
| 2500 | 40 | 11.2 |
Thus, we see that the magnitude of the “structural energy” of the molecules of living matter is indeed of the same order as the magnitude of the energy of activation. Of course, one cannot here expect an exact quantitative coincidence, since these figures themselves were obtained as average magnitudes independently of any definite reaction and its peculiarities, and also independently of the state of the living tissue, that is, of that changing distance at which its molecules are from the equilibrium position. From the fact that the structural energy of living molecules, calculated and measured in Hill’s experiments, although of the same order of magnitude as the energy of activation of chemical reactions proceeding in vitro, nonetheless exceeds the latter by 3–4 times, one can conclude that the degree of deformation of living molecules is higher than that which is, in general, called forth in the majority of reactions by the collisions of reacting molecules—a conclusion which already appeared probable from the very beginning.
To avoid misunderstandings, we must here emphasise still the following: when we have to do with reactions in vitro, in which the molecules entering into reaction (that is, the reaction tangle) receive their energy of activation only during the collision or through it, then a greater energy of activation signifies a smaller rate of reaction, and indeed even a very insignificant increase of the energy of activation calls forth a very considerable diminution of the rate of reaction. Were we simply to transfer this dependence to reactions proceeding in living tissue, then a strengthening of the energy of activation by 3–4 times would mean that there these reactions proceed infinitely slowly. However, such a transference would be incorrect, since this dependence is conditioned precisely by the fact that the molecules entering into reaction must be activated by means of collisions. Since the energy of activation which must be imparted to them is great, while the molecules entering into reaction are, so to speak, difficult to deform, the reaction proceeds slowly. If, however, the molecule is already in a non-equilibrium state and strongly deformed, as in living tissue, then of course such a dependence cannot hold.
Thus we have now obtained a conception of the course of the process of metabolism on the basis of that theory of living matter at which we arrived in this chapter, proceeding from the general laws of motion. This conception amounts in general outline to the following: a molecule of living matter is in a non-equilibrium state and therefore possesses unsaturated valencies. The processes of cleavage and combustion taking place in the organism occur precisely in consequence of this state of the molecules of living matter. The molecules of nutritive substances are bound by the free valencies of the molecules of living matter, being themselves deformable by the field of the latter upon collision with them. Thereby a reaction tangle is formed, in which a rebuilding takes place, the potential energy of the molecules of the nutritive substances going directly to prevent the transition of the molecule of living matter into the equilibrium state, or going to the restoration of this molecule after the rebuilding. But in view of the fact that, in the absence of growth, the products of cleavage of the molecules of the nutritive substances and the heat corresponding to this process of cleavage are liberated and given off—in view of this the arisen molecule would have to pass over again immediately into the equilibrium state, were there no metabolism, that is, were it not, even before the onset of this equilibrium, to collide with new molecules of nutritive substances, and so forth. Thus, the processes of metabolism supplying energy constantly maintain the molecule of living matter in its non-equilibrium state.
Depicting these processes schematically, we obtain the following scheme in the case of metabolism not accompanied by growth:
In this scheme E denotes a molecule of living matter, N the molecule of the nutritive substance to be assimilated, n₁, n₂, … the arising products of cleavage, and Q the evolution of heat corresponding to this process of cleavage. The designations enclosed in a circle denote the reaction tangles constantly arising upon repeated collisions with a molecule of the nutritive substance. The strokes attached to E denote the deformed active state of the molecules; the diminution of their number must mean that the distance at which the molecule of living matter is from the equilibrium state does not remain unchanged, but of necessity diminishes. This follows from the formula, deduced further on independently of the theory of living matter, according to which a diminution of free energy must take place at unchanged mass, that is, in the absence of growth (of increase of the mass of living matter). Consequently, this formula must retain its significance for the given special mechanism as well. It is, moreover, the direct result of the work of the structural forces and is equivalent to the conclusion at which Rubner arrives on the basis of his law of the dependence between the duration of life and the intensity of metabolism. This conclusion he formulates as follows: “The cleavage of organic nutritive products and the conversion of their energy bound up with this are accompanied also by constant changes in the grouping of the atoms of protoplasm, that is, so to speak, by work within and on the surface of living substance.”[Author’s note:] M. Rubner, Das Problem der Lebensdauer und seine Beziehungen, etc., München u. Berlin, p. 205, 1908. Work within and on the surface of living substance, accompanied by changes in the grouping of the atoms, can of course take place only when the structure of the molecules of living matter is in non-equilibrium, from which it further follows that the more cleavage takes place, the smaller the capacity for work that the structure will possess, that is, the molecules will approach equilibrium, as is depicted in our scheme by the diminution of the number of strokes; to the same conclusion Rubner too must have come.
If in the course of metabolism the indicated diminution of the free energy of the molecule of living matter did not take place, then the picture of metabolism given by us could simply be interpreted in such a way that the molecule plays for us the role of an ordinary catalyst and constantly comes out of the reaction in an unchanged form. This diminution of free energy is of course not an accidental or unimportant phenomenon: it is a necessary consequence of the fact that we here have to do precisely not with a catalytic action or a ferment in the generally accepted sense of the word. In the latter case the course of the reaction and the liberation of energy during it would not be a necessary condition of the existence of the ferment itself; in the former the dependence would be one-sided: the reaction cannot take place without the catalyst (or proceeds infinitely slowly), but the catalyst can exist even without the reaction which it accelerates. In our case, however, the matter stands thus, that the catalyst (that is, the deformed, non-equilibrium state of the molecule acting as a “catalyst”), on the one hand, itself participates in the reaction, and not merely transmits it, and, on the other hand, can exist only in so far and so long as it can preserve such a non-equilibrium state by virtue of the reactions which become possible in consequence of this state. That the matter really stands thus is again directly proved by facts. Thus, for example, we know that if one simply lets yeast zymase stand, then it “is inactivated,” but this inactivation does not set in, or sets in considerably later, if one adds grape sugar to the zymase and thus maintains the process of fermentation. We must, as a general rule, expect that if there are no molecules of nutritive substances which can be cleaved and rebuilt in the indicated manner upon collision with molecules of living matter, then the latter must pass over into the equilibrium state and lose their “enzymatic” capacity. Such a case we indeed have before us in the form of so-called autolysis. If we have a living tissue left to itself and to which no nutritive substances are supplied (that is, molecules with which it can react so as to ensure the maintenance of its state), then the molecules of this living tissue decompose. These phenomena of autolysis are usually explained thus: every living tissue, every living cell, also contains a proteolytic ferment. But then it remains incomprehensible why autolysis takes place only in dying tissue and not in living. Here again one tries to have recourse to the aid of hypothetical boundary surfaces and membranes within cells, supposedly hindering the ferments from coming into contact with the protein, or else one assumes that the conditions of the medium, such as acidity and the like, in the liquid phase hinder the action of the ferments in the living cell. The so-called latent period of autolysis is then explained by the fact that a certain time is required until, in consequence of dying, the conditions of the medium change to such a degree that the ferment is activated, or until the boundary surfaces disappear to such a degree that contact between the ferment and the protein becomes possible. However, we know from experience that if one adds gelatine, it is cleaved immediately, without any latent period; consequently, the ferment was active already from the very beginning. Thus, precisely in the phenomena of autolysis the conceptions of a special ferment, existing independently of the molecules out of which living matter is built and conditioning the decomposition of this matter—but only after its death—prove insufficient. Even if we wished to preserve this conception, or, more precisely, the terminology (since a concrete conception does not, after all, exist here), then even so all these facts would mean that what changes are not the conditions of action and the medium of the ferment itself, but the substrate subject to decomposition, that is, that the protein upon dying changes and becomes subject to the action of the ferment, in consequence of which autolysis sets in. Consequently, even adhering to the usual terminology, we are compelled to accept that the molecules of protein in life are in a different state than after death. On the other hand, we see that if we proceed from the fact of the deformed, non-equilibrium state of the molecule, directly explaining, as we have seen, their “enzymatic” character too, their chemically active state, then the supposition of the presence of a special proteolytic ferment, present in every cell but becoming active only after death, turns out to be superfluous. The immediate onset of the proteolysis of gelatine is then explained by the presence of a still active state in the molecules of the surviving tissue; the decomposition after the conclusion of the latent period is the result of the absence of reactions maintaining the non-equilibrium state of the molecules and conditioned precisely by this state; while the latent period itself is the consequence of the transition from the active, deformed state into the equilibrium one—a transition not yet accompanied by appreciable chemical processes and preceding the decomposition measurable by chemical methods.
It remains for us still to investigate the question of what chemical changes, upon this transition of the molecules into the equilibrium state, that is, upon dying, we are to expect on the basis of the theory of living matter developed here.
From the fact that the molecules of living matter, in consequence of their non-equilibrium structure, possess valencies disappearing upon the transition into the equilibrium state, it follows that the groups of atoms, or molecules, bound by these residual valencies, must, upon dying, be liberated even before the chemically registrable further decomposition of the molecules sets in.
We think that precisely by this is explained the “traumatic formation of ammonia,” investigated in detail by Parnas. Parnas[Author’s note:] Parnas u. Mozolowski, Bioch. Zschr., Bd. 184, 1927. showed that upon the mechanical destruction of tissue a rapid formation of ammonia takes place, proceeding within the course of one minute. Parnas emphasises that the intensified traumatic formation of ammonia is a special property of striated skeletal musculature, and adduces a table of such traumatic formation of ammonia for other tissues, from which it is evident that all tissues possess this capacity, though in smaller measure. Parnas also showed that blood plasma without blood corpuscles too forms ammonia, this formation taking place rapidly only in the first hours, but continuing, though in weakened degree, for 24 hours. If there are red blood corpuscles in the plasma, then an intensified formation of ammonia takes place upon their haemolysis.
It turned out that “the magnitude of the accumulation of traumatic ammonia represents the upper boundary of the physiological fluctuations of the content of ammonia,” and, further, that “rigor under the influence of heat leads to an accumulation of ammonia reaching the same height as the traumatic.” If we now take into account that, according to our principle, the muscle performs external work upon contraction at the cost of the structural forces, that is, that its structure then approaches the equilibrium state; further, that blood serum is spontaneously inactivated upon standing, the dielectric constant and surface tension diminishing in consequence of the transition of the molecules into the equilibrium state—then all these facts show that the traumatic formation of ammonia is a phenomenon accompanying the transition of the deformed molecules of living matter into the equilibrium state, and conditioned by this transition, in so far as certain NH₂ groups are bound in the living molecule by those residual valencies which disappear upon the transition into the equilibrium state. Especially in favour of this speaks the observation that the formation of ammonia upon autolysis is not subject to the law of mass action. There naturally arises the thought that the amino acids of the food proteins assimilated by living matter are, even before their transformation in the reaction tangle, bound by their NH₂ groups precisely by means of such residual valencies as give off the traumatic ammonia upon their disappearance in consequence of dying. We must also expect that the molecules of other assimilated nutritive substances too, for example the molecules of fats and carbohydrates, are at the first moment bound by certain residual valencies of the molecules of living matter even before their transformation, and are therefore liberated upon dying. A proof here could be furnished by the phenomena of the so-called separating-out of protoplasm upon autolysis and upon pathological, degenerative, or necrobiotic processes proceeding in vivo. As is known, in these cases we observe in the cell the liberation of fatty or lipoid granules, which until then were bound in one or another form and could not be detected by methods of staining. In the same way is explained the observation of du Noüy, who found that the addition of sodium oleate to blood serum at the first moment, it is true, considerably lowers the surface tension, but already after a few minutes the latter returns to the norm. This phenomenon can of course be explained only in such a way that the molecules of the added fat are bound by the protein molecules of the active serum. In favour of the view that the residual valencies making this binding possible exist precisely owing to the active state of these molecules, speak certain data according to which in inactive serum the return to the norm does not take place fully or even does not take place at all.[Author’s note:] Isar, Zschr. f. exp. Therapie und Immunolog. Forsch., Bd. 21; Blumenthal, Ibid.; L. Farmer-Loeb, Biochr. Zschr., Bd. 136, 1923.
Thus, the general laws of living systems set forth in the preceding chapters lead us with necessity to the conclusion that the non-equilibrium of living matter is explained not by the multiphase character and the constitution of the boundaries of phases, but by the molecular structure itself. We were able to show that this conclusion can be verified by means of physical and chemical methods of investigation, and that the facts (in so far as they exist) fully confirm it. Further, we showed that the theory of living matter based on these admissions and facts permits us to explain metabolism and its general regularities, and to form for ourselves a concrete conception of the mechanism of these processes. The conclusions drawn by us from the general laws of motion concerning the regularities of metabolism, the connection between structure and the processes supplying energy—these conclusions follow also from our theory of living matter. We have obtained a series of magnitudes proportionally to which the potential of living substance changes and by means of which it can, consequently, be measured. Such measures of the potential turned out to be: the quantity of heat liberated upon dying, the wavelength of the rays emitted, or not absorbed in life but absorbed after death, the electrical difference of potential between living and dead tissue, the dielectric constant, the surface tension, and so forth; all these magnitudes can be applied to the measurement of the potential, depending on the conditions of the experiment.
It remains to answer only one more question: can one, and in what respect can one, chemically determine the molecule of living matter in its deformed, non-equilibrium state? Have we here to do with protein molecules in the ordinary sense of the word? To this question it must be answered that the chemical determination of the molecules of solid phases in itself presents difficulties. These difficulties are still further increased for complex, micellar organic compounds. Thus, for example, K. Meyer writes: “Micellar compounds cannot be regarded as chemically homogeneous even when they are built out of simple constituent parts, for example out of glucose. To a still greater degree this holds for protein bodies. Concerning silk fibroin, usually called simply a protein body, the following can be said with certainty: this fibroin consists of micelles arranged in the form of a lattice, built out of simple molecules, but which nonetheless can as little be considered homogeneous as cellulose. In view of this it makes no sense to seek a ‘formula’ for ‘silk fibroin’ and to determine the magnitude of the molecules, for example, in such a way that one calculates the quantity of the product of decomposition occurring in the smallest amount, and then assumes that each ‘molecule’ must contain this product of decomposition at least in the form of one constituent part. The presence of an equally complex situation must be assumed for other protein bodies as well. We consider ourselves entitled to assert that practically there exists in general no ‘single’ protein body.”[Author’s note:] K. Meyer, Bioch. Zschr., Bd. 208, 1921. If, then, the chemical determination of these compounds presents such difficulties even in the equilibrium state outside the organism, then the more is this the case in the living state. Upon consideration of the above-adduced scheme of metabolism the question arises: which moment do we, properly speaking, consider? The moment E, or the moment δ₁N₁? And can one in general grasp the moment δ₁, that is, the molecule not in metabolism and not in the process of rebuilding? At the moment of metabolism the molecule, as we have seen, is so closely bound up with carbohydrates, lipoids, amino acids, and so forth, that we have no right to consider it apart from them. If in the protein bodies passing out of living tissues we never obtain a chemically single body, and there are moreover always micelles present, then these facts are explained precisely by this circumstance and by the conception of metabolism proposed by us. It is therefore understandable that many physiologists and biochemists refuse to speak of protein as the “bearer of life” and attach the same significance to lipoids, salts, and so forth. But, as we have already shown, such a conception entails a fundamentally false picture, if one does not admit a special non-equilibrium state in the molecular structure. If, however, one admits a special state of the molecular structure—and this is dictated to us by the regularities of the biological order—then in this state there exist not all kinds of molecules; and if they temporarily exist in it, then in consequence of this state of the basic building materials of living matter, which, although not identical with the protein bodies chemically obtained by us from organisms, nonetheless upon dying decompose into the building materials of this protein. Therefore we are entitled and even obliged, in the sense of the ideas set forth in the present chapter, to speak of “living protein.”
Part II: Theory of Vital Phenomena
Introduction
In this part we shall attempt to give, in so far as this is possible, a theory of those vital phenomena which — such as metabolism, growth, irritability, reproduction, heredity and evolution — are common to all living beings, and we shall build this theory upon the basis of the general laws of motion and the theory of living matter set forth in the first part. The vital phenomena just named are usually designated as the necessary marks, as the criteria of life, or of living systems. Before we investigate whether the elaboration of a theory of these phenomena on the basis of the general laws of motion and the theory of living matter is at all possible, we must make clear to ourselves what the interrelations are between these phenomena and the general laws of motion of living matter. If these phenomena are indeed necessary marks, necessary manifestations of every living system, then this means that a system deprived of one or several of these marks cannot be regarded as living. But since we designate as living all material systems that satisfy the laws of motion set forth, this means that all these marks must already be contained in the general laws of motion of living matter, that is, they must of necessity follow from the latter or be derivable from them. In other words, if we regard the general vital phenomena just named as necessary criteria of life, then we may speak of general laws of living matter only in the case where these criteria are already contained within them. Accordingly, we must then see one of the conditions for the correctness of any general laws of motion in the possibility of deriving from them these general vital phenomena. Consequently, in the case of our laws of motion too, we should have to prove here that these phenomena follow from them of necessity. The proof of this necessity would itself constitute the theory of the phenomena in question, in so far as we should succeed in showing that the latter find their explanation on the basis of that form of motion, peculiar to living matter alone, which is expressed by our laws. Were this to prove impossible, we should have to conclude that our general laws do not express that form of motion which is peculiar to living matter and to it alone. From the outset, however, it is clear to us that at least some of these vital phenomena, as for example reproduction, are not necessary in order to call a given system living: for example, we may withhold fertilisation in some animal with sexual reproduction; it will then not reproduce, but it will nevertheless still live for some time, we shall call it living, and our general laws of motion of living matter will retain their validity for it. The matter stands more intricately with irritability. But in any case, here too we can, at least theoretically, conceive that a living being is held under such conditions that no stimulation acts upon it, that is, no harmful influence sufficient to bring about the damage to structure that produces external work. We have already said (see Chapter II) that for the reception of food external work is necessary, that is, stimulations. Therefore a living being under such conditions will starve and gradually die. But it will nevertheless live for some time, and our laws of motion will retain their validity for it, although one of the criteria of life — irritability, that is, the phenomena of stimulation — will not be manifested. We can go still further and conceive a living system completely isolated from those substances which can serve it as food, that is, we conceive a cell in which there are no reserve substances capable of serving as a source of energy for its metabolism. Such a system will perish very quickly, since it cannot use the energy of food molecules to maintain the non-equilibrium state of the molecules of living matter
according to the scheme in Chapter IV. Consequently, in such a system metabolism will be absent. The question arises whether in this case our general laws of motion do not lose their validity, and whether we may still, under such conditions, call this system living? If we answer this question in the affirmative, then it will turn out that metabolism too is not necessary in order to call a system living. In this case, however, this answer is not given to us so easily as in the investigation of the role of reproduction and irritability, and with respect to metabolism the question stands in need of a more detailed analysis. In essence, the matter here concerns the following: if we have a system consisting only of living molecules , which have no possibility of binding and activating molecules of food by means of the free valences that these living molecules acquire owing to their non-equilibrium structure, and of using their energy to retard equalisation or to restore the non-equilibrium state, then how, in such a case, will the processes of equalisation proceed: will work be performed in this case against the expected equilibrium at the expense of the energy released in such equalisation, or not? In other words, will equilibrium set in, in this case, in the same way as takes place under the given conditions according to the laws of physics and chemistry, that is, without internal work of the systemic forces directed against equilibrium? This question is equivalent to the question whether in this case too there exists a difference between the two integrals
(see Chapter I, p. 33) or not. For it is entirely clear that if we kill some animal or cell, then the reserve substances contained in them (carbohydrates, proteins, fats, etc.) are not used, whereas in the spontaneous dying of an animal they are at least partially used, the free energy in the latter case being less than in the former, so that, owing to this difference, our two integrals too may turn out to be different. How, then, does the matter stand if there are no such substances, decomposable and assimilable by the molecules of living matter, that is, if metabolism is impossible in the sense of the use of chemical energy or the assimilation of molecules not belonging to living matter? It is clear that if we have only one molecule of living matter, that is, only one living protein molecule in the above-mentioned sense (see Chapter IV), then this molecule will be unable to perform work against equilibrium at the expense of the energy it possesses owing to its deformed state. It cannot maintain itself in this state or retard equalisation. Consequently, a single molecule, isolated from other molecules which it might assimilate, is not in a condition to live. But we can conceive that if there is a large number, several thousand molecules of living matter, then even without the assimilation of other molecules the energy peculiar to one part of these living molecules in consequence of their non-equilibrium state may serve either to maintain the non-equilibrium state of others or to retard in the latter the processes of equalisation. We know that absolutely starving organisms do not die immediately after they have used up all the assimilable reserve substances contained in them, but use part of their own living protoplasm. Thus, for example, planarians, when starving, are able to use up the greater part of their body and to diminish manyfold. Consequently, in this case the starving animal lives without metabolism, that is, without the assimilation of non-living molecules. One might, it is true, say that in this case too metabolism takes place, in so far as here living molecules belonging to the system are assimilated by other living molecules belonging to the same system. Here there is no activation of a molecule not belonging to the system, nor its rebuilding and incorporation into the number of the system’s molecules, just as here the energy of such a molecule is not used to maintain the non-equilibrium state; but here we are dealing with an interaction between molecules of the living system, the internal work being performed at the expense of the energy of molecules already active, that is, at the expense of the structural energy already present in consequence of the non-equilibrium state. Since in this case no work is needed to bring the molecules into the non-equilibrium state, and the energy of these molecules is used directly for work against the onset of equilibrium in other molecules, it follows from this that in the “assimilation” of such living protein molecules by other living protein molecules of the same system the potential of living matter, that is, the average distance of the molecules from the equilibrium state, must not diminish, but on the contrary may in this case increase at the expense of part of the living mass. We have here before us a process essentially different from metabolism. This process plays an outstanding biological role, especially in reproduction (see Chapter II).
We thus see that a system displaying no metabolism too can for some time be called living, and in so far as this system may possess the form of motion expressed by our laws, to that extent it must be called living. Of course, in the case of absolute starvation this system must inevitably die after a definite time, just as is the case in the absence of stimulations and of reproduction.
We thus arrive at the conclusion that the general vital phenomena just named, such as metabolism, growth, irritability, reproduction, are not necessary marks of life, that is, do not serve as its criteria. Consequently, they are not contained in the form of motion of living matter and cannot be derived from its general laws of motion.
All these phenomena possess, however, the following common mark: they are all necessary conditions for the constant viability of living matter, that is, for the maintenance of life. In chemistry and physics we are accustomed to understand by the necessary conditions for the existence of some substance or state only certain boundary conditions, lying outside the system and reproducible independently of it, such as temperature, pressure, light or some other influx of energy or matter. This is understandable, since here we are dealing with the existence of equilibrium states, which is determined by the relation between the phases entering into these states and the boundary conditions, and in so far as these states are equilibrium states, they will exist constantly, so long as the boundary conditions are maintained. If we have a definite quantity of gas at a definite temperature and a definite pressure, then no question whatsoever arises regarding the capacity for existence of this gas: it will exist as a gas and display the laws of motion peculiar to the gaseous state, until we alter the conditions of temperature and pressure. And here there is no need for any special manifestation of the gas laws in order to maintain this state; it is maintained by the conditions themselves, since here we are indeed dealing with equilibrium. The necessary conditions for the constant existence or maintenance of some state in inanimate nature are, consequently, always conditions of the surrounding sphere, which to this end must be and can be created, and not manifestations of the system itself. We can also indicate definite limits of these external conditions; for example, for isolated gases, and therefore gases incapable of entering into chemical reactions with other substances, we can indicate exactly that critical temperature above which they always remain in the gaseous state, that is, follow the gas laws; for example, with respect to CO2 we can say that it will always exist only in the form of a gas, so long as the temperature is above 32° C.
The question of the conditions necessary for lasting viability, that is, for the preservation of living matter, is an altogether different one, since here we are dealing with the lasting existence of a non-equilibrium state. Therefore here we are dealing not with conditions of the environment independent of the laws of motion of the given system or the given state of matter, but with such as are a certain expression of these very forms of motion, of the very state of living matter. Therefore, in order to avoid a confusion of concepts, one should not say that the mentioned vital phenomena constitute necessary conditions for the lasting viability or preservation of living matter, but should rather regard them as those particular manifestations of the form of motion of living matter which have led to its preservation on earth. If such manifestations of the form of motion of living matter as lead to its preservation are to be designated as adaptations, then all the mentioned general vital phenomena must be regarded not as criteria or necessary marks of life, that is, of the form of motion of living matter, but as adaptations. But since these adaptations are particular manifestations of the form of motion of living matter, and the form of motion is expressed by our general laws of motion, it follows from this that the possibility of these manifestations (adaptations) must be derivable from our general laws of motion of living matter. Consequently, the form of motion of living matter contains within itself these general vital phenomena as possibilities. The necessity, however, of the realisation of these possibilities does not follow from the laws of motion of living matter. Their realisation depends, of course, also on the conditions of the environment. The realisation of the possibility of metabolism, contained in the form of motion of living matter, in a definite living system depends, of course, essentially on the conditions of the surrounding environment, namely on whether there are present in the surrounding environment molecules assimilable by the given system, and, for green plants, for example, light as well. This dependence of the realisation of the possibilities given by the form of motion of living matter upon the external environment is evident also for any other vital phenomenon, for any given living system. Therefore we can indicate for each separate living being, at least theoretically, those critical values of the external environment beyond which it is no longer capable of lasting existence and must die after a definite time, that is, the living matter of the given system must pass over into a non-living state possessing other laws of motion. In the same way we can indicate, for example, the critical temperature of any liquid, that is, the temperature above which the liquid is absolutely incapable of continuing to exist as such and must pass over into another state, the gaseous, possessing other laws of motion. We shall call these limits the “limits of physiological adaptation.” But already here we meet an essential difference in comparison with inanimate matter. Namely, whereas an inanimate system within these critical limits is capable of constant existence, a living system, even within these limits, that is, the limits of physiological adaptation, likewise cannot exist constantly, that is, unlimitedly in time. A living system acquires the capacity for constant existence only through reproduction. Theoretically, reproduction could ensure the constant viability of living matter, that is, of a given kind of living system, on condition that the sources of energy in the surrounding environment, or the space in which they exist and in which the external conditions do not pass beyond the limits critical for these systems, may be regarded as infinite. But in actual fact, as we know, this never takes place, and living systems can preserve constant viability even beyond the limits of physiological adaptability by means of hereditary variability, which leads to the emergence of such systems as become constantly viable even beyond the external conditions critical for their ancestors. Thus hereditary variability too belongs among those universal vital phenomena which have led to the preservation of living matter on earth and are a necessary condition for its constant preservation; therefore it too can be designated as an adaptation. Consequently, its possibility too must be contained in the form of motion of living matter as a particular manifestation of its general laws of motion. Thus living matter, owing to the form of motion peculiar to it, can push back the limits set to its constant viability by external conditions. Therefore we can in principle indicate the limits of constant viability for each existing form of living matter, but, at least at the present moment, we cannot indicate them for living matter in general. We know only that at the present time living matter is viable under all external conditions existing on earth: in water, in earth, in air, with oxygen and without it, at enormous hydrostatic pressures, on the sea bottom and at the weak atmospheric pressures in the upper layers of the air; at temperatures above 80° C, where every ferment and every protein body outside living matter is non-viable and coagulates. It is equally difficult for us to indicate the lowest temperature limit, since organisms are known to us that withstand the temperature of liquid helium without losing their viability. Therefore we cannot, for example, indicate to what temperature the earth must cool down in order to exclude the possibility of the existence of living matter. All these hereditary variations of living matter are the realisation of certain special manifestations of the form of motion of living matter, possibilities contained in this form of motion and realised in the course of evolution, just like the remaining general vital phenomena which, as we have seen, are wrongly called criteria or necessary marks of living matter, such as: metabolism, reproduction, irritability, etc. Therefore their
necessity cannot be derived from the form of motion of living matter, that is, from the general laws of motion expressing this form. But we must be in a position to show that they are possibilities contained in this form of motion, and that they really constitute its special manifestations, that is, particular cases of our general laws of motion. To derive the necessity of these phenomena from our laws would be possible only in the case where we assumed that all the particular manifestations of the form of motion of living matter, that is, all the possibilities contained in it, are of necessity realised in the course of evolution. But even then they would not be criteria of life, or of living matter. There exists only one criterion of living matter: its form of motion, which finds its expression in certain general laws of motion of living matter.
Chapter 1: Metabolism and the Boundary of Assimilation
The general laws of living matter set forth in the preceding chapter lead us directly to definite conceptions concerning metabolism [обмен веществ — obmen veshchestv] in the organism and already permit us now to deduce certain general regularities governing it.Обмен веществ — literally “exchange of substances.” We render it throughout by the established term “metabolism,” but flag the literal sense here at the outset, since Bauer’s energetic argument leans on the image of substances being taken up, transformed, and broken down.
First of all, it is clear that the general laws set forth above contain metabolism within themselves as a general property of every living system. Indeed, if a living system is not in equilibrium, and if this non-equilibrium state is stable thanks to the fact that it is maintained by the free energy of the system itself at the cost of the constant processes of equalisation [выравнивание — vyravnivanie] proceeding within the latter—if all this is so, then it can take place only in the case where the internal conditions of the system (that is, the structure of living matter) themselves change and thereby are constantly renewed and restored. Consequently, the system can in no way be materially closed. One cannot return to one and the same matter its former non-equilibrium state with its former free energy after equalisation has taken place, that is, after the diminution of its free energy. This would contradict the second principle of thermodynamics if the system were in general isolated, and it would equally contradict it if the system were constantly brought into a new state capable of work only at the cost of thermal energy supplied from without. Just as little can this take place at the cost of work performed from without against the system, since this would mean that the source of the work lies outside the system, that is, the presence of a working system bound to the living system would be presupposed. Thus, the non-equilibrium state can be maintained at the cost of the free energy of the system only on condition that the system is not closed and that the substances taken up are directly brought into the non-equilibrium state proper to the structure. The work necessary for this is performed at the cost of the energy of the system’s processes of equalisation, that is, at the cost of structural energy. The chemical energy of the food substances taken up is consequently used only in a roundabout way, after they have been brought into the non-equilibrium state of the structure of the living system upon their equalisation, that is, their destruction. The building-up of the living structure, that is, the bringing of the substances taken up into the state proper to the matter of the living system, we call assimilation [ассимиляция — assimiliatsiia]. The process of equalisation that accompanies the destruction of this structure, in which is liberated the energy necessary for bringing the substances taken up into the non-equilibrium state of the structure of the living system, we call dissimilation [диссимиляция — dissimiliatsiia]. It is clear that when assimilation prevails over dissimilation, that is, when more substance passes into the non-equilibrium state of living matter than is annihilated by the latter, then we have to do with an increase in the quantity of living mass, of living matter, that is, we have to do with growth.
We shall now occupy ourselves with the question of whether our general laws of motion of living systems permit us to draw more precise conclusions about the relations in which assimilation and dissimilation must stand to one another, and about the conditions under which the one or the other will predominate.
But first let us point to the following circumstances.
Metabolism, that is, the uptake of substances from the surrounding environment, the restoration and decay of living substance, is a general property of all living systems. In the laws of motion, of stable non-equilibrium, and of the work of the systemic conditions set forth by us, this property is contained as a possibility. It is, as we have seen, a necessary consequence of these laws, but not conversely; it is an expression of them, in so far as the living system is preserved over time. Thus, our general laws of motion contain within them more than the necessity of metabolism with dissimilation and assimilation. From them one can, over and above this, deduce something also about the paths and the mutual conditioning of these processes, and further, as we shall presently see, about their relations to one another. It would therefore be an error to hold that metabolism, assimilation, and dissimilation themselves already contain the laws of motion, or still more the characteristic form of motion of living matter. A detailed investigation of the forms and the course of these processes of exchange would, of course, also lead us to the general laws of motion. But the same then applies also to every general property, such as irritability, reproduction, growth, and so forth. A deeper penetration into the course of these processes would give us an ever more precise conception concerning the state, the structure of living substance, and on the basis of this conception we should arrive at the general laws. But our general laws contain something more as well, precisely in the sense that they already permit certain conclusions also concerning the structure and state of living substance. And these conclusions were obtained not only by way of generalisation on the basis of the phenomena of metabolism, nor indeed could they have been obtained by such a path. We obtained them, as indicated in Chapter I, by means of bringing together and abstraction from the regularities of the most diverse properties of living beings. They therefore also permit certain more precise general conclusions concerning metabolism and its regularities.
Finally, it must still be pointed out that from our laws of motion there follows not only the necessity (in the indicated sense) of metabolism, assimilation, and dissimilation, but from them one can also draw a conclusion concerning the form of these processes as well—a conclusion contradicting the generally widespread conceptions, especially those set forth in textbooks.
We have in mind the division into so-called plastic and dynamic, or energetic, metabolism. Generally speaking, biologists and physiologists still represent the matter to themselves in such a way that certain nutrient substances, or their chemical energy liberated upon the combustion or enzymatic splitting of these substances in the organism, serve as the direct source of work, while other substances (or under certain conditions the same ones) are used for the building-up or replacement of the structures of the living system. In the first case one speaks of energetic or dynamic, in the second of plastic metabolism. From our principle of the work of the structural forces, coinciding in essence, as we have seen, with the principle of stable non-equilibrium, it follows, however, that this division is incorrect and that, in essence, there exists only one plastic metabolism. For our principle asserts that the chemical energy of the food taken up is converted into work not directly, but only in a roundabout way, through its passage into the state and structure of the living system and its constituent parts. It further asserts that external work, that is, work in the sense of physics, can be performed only at the cost of the energy of this structure by way of its destruction, as a result of the processes of equalisation, in the case where, in consequence of some disturbance, this energy of dissimilation cannot be employed for assimilation. There are no energy-supplying processes, such as for example glycolysis, the oxidation of fats, and so forth, that would proceed independently of the constitution and state of the matter of living systems. The energy of these processes is liberated only after the substances have passed into the state and structure of living matter, which precisely determine the non-equilibrium state, that is, the capacity for work. It is easy to see that the conception of an energetic metabolism independent of the plastic is rooted, firstly, in an erroneous conception of the state of living structures as one of equilibrium, and secondly, in the growing significance of external work, as was set forth in the preceding section. This conception holds very firmly, despite the conclusion at which all biologists occupied with the thermodynamics of vital phenomena have already arrived—that the energy-supplying processes are needed for the preservation of certain non-equilibrium states (for example, Warburg, Lipschitz). It holds also despite the opinion, already expressed and substantiated by many physiologists, that it is the tissue itself that is dissimilated, and not the substances taken up into the blood or entering the cell. Richet already arrived at this conclusion on the basis of his experiments on the course of the temperature curve after death. Besides those indicated, there are still the following causes why this incorrect conception is yet preserved: by non-equilibrium states one always understood only such as are conditioned by definite properties of the system, for example differences of concentration conditioned by the semipermeable properties of the walls; in the best case it was thought that these properties of the system are preserved constant thanks to a supply of energy from without, as in a dynamic equilibrium, and in this way the possibility was obtained of directly applying the principles of thermodynamics. But no one thought that the conditions of the system themselves change and that the direct source of energy for the preservation of non-equilibrium is precisely these changes, and not the equalisation, for example, of differences of concentration, that is, of differences of thermodynamic potential of the various phases conditioned by the conditions of the system.
Another cause was and remains the fact that we can isolate from cells enzymes or substances containing enzymes that evoke the given energy-supplying processes also in vitro, that is, independently of any structure whatever. As we have already briefly indicated in the chapter on the theory of living matter, enzymatic action too is bound up with an excited, deformed state of the molecules. Here we shall only point out that all the facts known to us concerning enzymes and the conditions of their action likewise speak rather in favour of the view that either the enzymes themselves possess non-equilibrium structures, or that their action is bound up with such structures. That their arising is bound up with the activity of living structures is generally known. Further, we know that all the enzymes known to us that play a role in organic metabolism, such as the proteolytic, glycolytic, and lipolytic enzymes, are spontaneously inactivated after isolation in vitro. We also know that enzymes lose their activity if a certain degree of their purity is exceeded. All these are known facts, speaking for the view that the arising and action of enzymes is conditioned by certain structures of the molecules or groups of atoms that form or carry them, these structures, moreover, not being preserved under unchanged conditions, that is, not being in equilibrium. Here too it is not difficult to notice that the explanation of the processes of metabolism by means of enzymes acting analogously to catalysts and essentially independent of the molecules out of which living substance is built—that this explanation proceeds from the tendency to explain the phenomena of metabolism by means of the unchanging structure of the particles of living systems and is likewise rooted in the incorrect conception of the state of living structures.
To illustrate that a deeper penetration into the processes of metabolism and of enzymatic reactions also leads us to conceptions concerning the course of metabolism and concerning the deformed state of molecular structures similar to those that were developed by us in the chapter on the theory of living matter, we shall adduce here certain considerations of Weichherz and Nord[Author’s note:] J. Weichherz and F. Nord, Kinetische und Molekuldynamische Betrachtungen zum Gärungsproblem, Protoplasma, Bd. X, 1930. from their investigations into the mechanism of action of enzymes, in particular into the process of fermentation: “Hitherto it has been possible to accept that the enzymatic reaction is a heterogeneous-catalytic reaction, that is, a surface reaction—we shall not, moreover, touch upon the question of whether the enzymes themselves possess a surface, or whether they are merely situated on a surface in an adsorbed state, since this question has no significance for our further considerations. But it is highly probable that the substrate subject to catalysis, that is, the sugar, must be adsorbed superficially thanks to the active centres situated on it, the carbon chains being, in consequence of the adsorption forces, deformed, that is, the relative position of the atoms or of their groups in the adsorbed state is other than in the free molecule of the solution. Thereby there is created such a molecular construction as could be designated the ‘fermentable’ form of the sugar, we should not, however, have the right to speak of a spatially isomeric transformation in the sense of classical structural chemistry. In consequence of the deformation of the molecule and the energetic activation of the valencies by the active centres, the internal energetic conditions in the sugar molecule are disturbed, the activated kinetic energy of the atoms in certain places exceeding the binding energy existing between them, in consequence of which the molecule disintegrates at this place. In this way there are formed fragments that are not, however, molecules in the classical sense, since they need not necessarily be closed off outwardly to the point of stability. This phase can be designated as the decomposition of the C₆ molecule into two labile C₃ molecules, and the theory of the sequence of phases of Wohl–Neuberger–
Neuberg–Kerb accepts at this place the formation of two molecules of methylglyoxal. But since the non-physiological methylglyoxal added does not split under the action of yeast, attempts were made to escape from this difficulty by asserting that what is accepted is the formation not of genuine methylglyoxal, but of some hydrogenated form of it or of an isomeric compound, which can then be converted into pyruvic acid.
This reasoning, founded only on formulae, could deceive concerning its weak sides only so long as the investigators operated with chemically precisely defined compounds or molecules of classical structural chemistry, reflecting only approximately the true grouping of atoms. But as soon as we no longer presuppose the arising of these energetically closed molecules, but deduce all further processes from the existence of labile fragments not yet stabilised in respect of chemical valency, we must regard the sequence of phases of Wohl–Neuberger–Neuberg–Kerb only as a translation of really existing relations into the language of structural chemistry, insufficient for the depiction of these processes. Thus, instead of the conceptions of structural chemistry, we here propose a ‘molecular-dynamic’ approach, which is as yet only qualitative. But only this molecular-dynamic approach gives us the possibility of explaining the course of fermentation, which has become more comprehensible to us in recent years thanks to the works of many authors.”
We have adduced this passage in full in order to show that a more detailed investigation of the enzymatic processes of metabolism compels us ever more to accept the existence of molecules that are not closed off in respect of chemical valency, but are excited, deformed, thus gradually bringing us closer to that conception concerning metabolism which we developed on the basis of our general laws of motion in the chapter on the theory of living matter. And in this domain too we shall be able to attain greater clarity by studying this state more deeply on the basis of our general laws.
After these remarks we shall occupy ourselves with the consideration of the question of what conclusions concerning the relations between assimilation and dissimilation in living systems we can draw from our general laws.
Let us first consider the process of growth of the living mass. This mass is a system whose free energy can diminish under the existing conditions, since the system is not in equilibrium.
As follows from our theory of living matter (see Chapter IV), this non-equilibrium is in the last analysis conditioned by the molecular state of living matter itself. When equilibrium has set in, that is, when the molecules pass into a stable state with the lowest level of energy under the given conditions, then we can no longer designate this mass as living.
The excess of free energy that arises in the living system in consequence of the excited, non-equilibrium state of its molecules in comparison with the unexcited equilibrium state, we shall in what follows briefly call the free energy of the living system, designating it by . If denotes the mass, and the free energy of unit living mass, that is, the thermodynamic potential [потенциал — potentsial] in the above-indicated sense, then of course:
But it is perfectly clear that if the system were closed, and the conditions (such as pressure, temperature) remained unchanged, then a diminution of would have to set in. This diminution takes place, firstly, in consequence of the lowering of the free energy of the excited molecules, that is, the approach of the latter to the equilibrium state, and secondly, in consequence of the diminution of the living mass itself, since we have indeed said that a mass whose molecules have passed into the equilibrium state can no longer be designated as living. An obstacle to such a process of equalisation can therefore be only the fact that the system is not closed, and that the mass which has attained the minimum of free energy, that is, which has died and become incapable of work, is replaced by “living mass” in the above-indicated precise sense. Such and only such an increase of the living mass, that is, of the mass of the protoplast in the non-equilibrium state in which it finds itself, we call assimilation. The synthesis and deposition of starch in plant cells, or of fat from food in animal tissues and cells, cannot consequently be considered as assimilation in the narrow sense, that is, as an increase of the mass of living substance. Only mass assimilated in the strict sense of the word, that is, only that entering into the phases of the protoplast with the same magnitudes of state and the same molecular structures as those conditioning the non-equilibrium state of the latter—only such mass is capable of work and yields, upon equalisation, energy serving for the performance of the vital functions. These processes of equalisation we call dissimilation. It is consequently the living substance, the protoplast itself, that is dissimilated. The chemical energy of the food taken up becomes suitable for vital manifestations only after or by virtue of its assimilation. This follows from our principle, according to which the direct source of all work of living systems is given in the processes of equalisation taking place within them. On the other hand, this source of energy, formed by dissimilation, must be employed for the work that leads to the establishment of the non-equilibrium state, to the formation of new living mass capable of work, that is, to assimilation. This requirement answers also to that schematic picture of metabolism which we gave in Chapter IV (p. 75). Since the system is not closed, and since we assert nothing either concerning the chemical potential of the mass taken up or concerning the degree of decay in the dissimilatory processes of equalisation and the level of the energy liberated thereby, we can of course, without introducing further restrictions, say nothing about whether the processes of increase of living mass, that is, assimilation, or the processes of equalisation within it, that is, dissimilation, must outweigh, or whether the one and the other must be in equilibrium. The relation of the energy liberated in dissimilation to the work necessary for the assimilation of the mass taken up, that is, for the passage of the latter into the given non-equilibrium state of the protoplast, depends, however, not so much on the heat of combustion or of splitting of the substances taken up as on the state and the structural conditions into which they fall upon assimilation, since the chemical energy of the food taken up becomes suitable for employment precisely only as a result of assimilation, while the energy of dissimilation must be employed precisely for bringing the food into the corresponding non-equilibrium state, that is, for the creation of living substance. Both processes are thus essentially connected as two sides of one and the same process. As to what quantitative relations exist between them, and by what these relations are regulated, we can say something only on the basis of general biological regularities. From our general laws of motion and from the theory of living matter developed in Chapter IV it follows that for the assimilation of a certain mass, that is, for the increase of living mass, a certain work of the already existing living substance is required, that is, a certain lowering of the potential ; therefore the first question we must put to ourselves is the following: how great is the lowering of the free energy of unit living mass, that is, , when unit mass is assimilated. This lowering will depend on the state of the already existing living mass, since the molecules of the food must indeed be brought into that state in which the molecules of the living mass precisely find themselves. If the potential of the latter is high, then more work will also be needed to raise the molecules to this level of energy; consequently, upon the assimilation of unit mass there will occur a greater fall of potential. Thus, the fall of potential upon the assimilation of unit mass will be proportional to the potential of the existing assimilating living mass, that is,
If we denote the initial mass by , and the initial potential by , then we obtain by integration:
Substituting this expression into formula (1), we obtain the free energy of the living system as a function of the living mass:
This function has the derivative:
Since the factor is always positive, then, so long as , there will take place an increase of the free energy of the system as a whole with the increase of its mass, up to a maximum lying at . The free energy of the system, with an increase of mass above this value, will fall.
In Fig. 3 is depicted the course of this function for various values of , and being moreover taken as equal to 1.
The value of is determined from formula (2). It is that fractional magnitude by which diminishes upon the assimilation of unit mass.
The magnitude , as is evident from formulae (2) and (4), has the dimension . If we denote by the magnitude of the mass at which the maximum of free energy is attained, then, since , we obtain:
Thus, up to this value growth, although it will lead to a diminution of the potential , will be accompanied by an increase of the entire free energy of the living system. The work which the structural forces thus perform upon assimilation is accompanied, up to this limit, by an increase of the capacity for work of the system in accordance with our principle of stable non-equilibrium. Were growth to continue even after the attainment of this boundary, this would lead to a lowering of the free energy of the system, but this would contradict our principle of stable non-equilibrium. Consequently, after the attainment of the value , the growth of living matter must cease. Hence that part of the curve (Fig. 3, curve I) which extends above this value is biologically not real. We have therefore depicted this descending branch of the curve in the figure by a dotted line. We call the boundary of growth, or the boundary of assimilation.
The question now arises: what happens after the attainment of this boundary of assimilation? As is evident from our scheme of metabolism on p. 75, the active, deformed state of the molecules of living matter cannot be preserved without metabolism. At the cost of the processes of equalisation the molecules of the food are activated, and their energy will serve for the maintenance of this state. Consequently, assimilation and dissimilation will continue. But if, after the attainment of the boundary of assimilation, the increase of living mass were to continue, then, as is evident from the above and from Fig. 3, this would lead to a diminution of free energy. Just as much would a diminution of mass, that is, a preponderance of dissimilation over assimilation, lead to a diminution of free energy. Thus, from our general laws of motion it follows that after the attainment of the boundary of assimilation a stationary state must set in with respect to the living mass. Dissimilation and assimilation must balance one another. In the thermodynamic respect too this state differs from the rest. Indeed, since the free energy of the living system as a function of mass possesses its maximum at the boundary of assimilation, that is, , the free energy, with continuing stationary metabolism, remains unchanged or exhibits only imperceptible fluctuations in comparison with those which would take place in the case of stationary metabolism before the attainment of the boundary of assimilation; that is, at the boundary of assimilation the state can be preserved by means of stationary metabolism with the least expenditure of work.
Thus, we arrive at the conclusion that both the initial preponderance of assimilation over dissimilation, and the existence of the boundary of assimilation, the setting-in of a stationary state with respect to the dissimilation–assimilation of living matter at this boundary, follow directly from our general laws of motion and from the theory of living matter. Let us emphasise once again that the stationary state with respect to mass at the boundary of assimilation has nothing in common with a dynamic equilibrium, since here this state is indeed created or preserved thanks to the constant work of the system itself, and not of any source of energy lying outside this system.
The dependence of free energy on mass obtained by us does not depend on the course in time, that is, on the speed of growth, since according to our formula (1) the fall of potential corresponding to the increase of mass depends only on the potential, that is, on the state of the assimilating mass, and does not depend on how quickly this state is attained. On the contrary, after the attainment of the boundary of assimilation, where with respect to mass a stationary state sets in, that is, where , the free energy of the living system becomes exclusively a function of time.
We shall now occupy ourselves with the investigation of the dependence of the free energy of the living system on time after the attainment of the boundary of assimilation. Can this state be preserved constantly, or must a diminution of free energy with time necessarily occur? In the ordinary language of biology this question means: is ageing a necessity, or does there exist in principle the possibility of individual immortality? A detailed investigation of the question of so-called “individual immortality” we shall find in the chapter on reproduction.
In the chapter on the theory of living matter we gave the scheme of metabolism precisely for the case where no growth takes place. Already from this scheme it follows that the potential of the living mass necessarily diminishes in this case too, but since in this case the mass remains constant, and since according to our definition the free energy of the living system is expressed by the product , it follows from this that the free energy of the system as a whole must also diminish. We shall now investigate in what manner this diminution with time can be expressed, and on what factors it depends.
If we had to do with a non-living system in a non-equilibrium state, then its free energy would diminish with time, this diminution taking place approximately, as we see in the majority of processes of equalisation, according to the formula:
But according to the principle of stable non-equilibrium the living system, at the cost of its structural forces, performs work against this equalisation; therefore here, as we have indicated, the initial conditions themselves do not remain constant, but will depend on the work of the structural forces at each moment, the free energy at each moment being proportional to the work performed by the system, that is, by its structural forces. We must consequently take as equal to , where is a factor of proportionality. The minus sign means that the work takes place at the cost of the free energy of the system itself and that thus this free energy must diminish.
Thus, for the free energy of the living system as a function of time after the attainment of the boundary of assimilation we obtain:
The solution of this differential equation gives:
This is a monotonically decreasing curve which, with the increase of , asymptotically approaches zero.As printed in the 1935 original, equation (6) carries the exponent , whereas solving equation (5) yields instead a form — an inconsistency between (5) and (6) in the source, reproduced here unaltered. In Fig. 4 is depicted the course of this curve for various values of with
unchanged (with )The capital here is the source’s misprint for the decay constant of equations (5)–(6); it is not the heat-proportionality constant introduced below. and for .
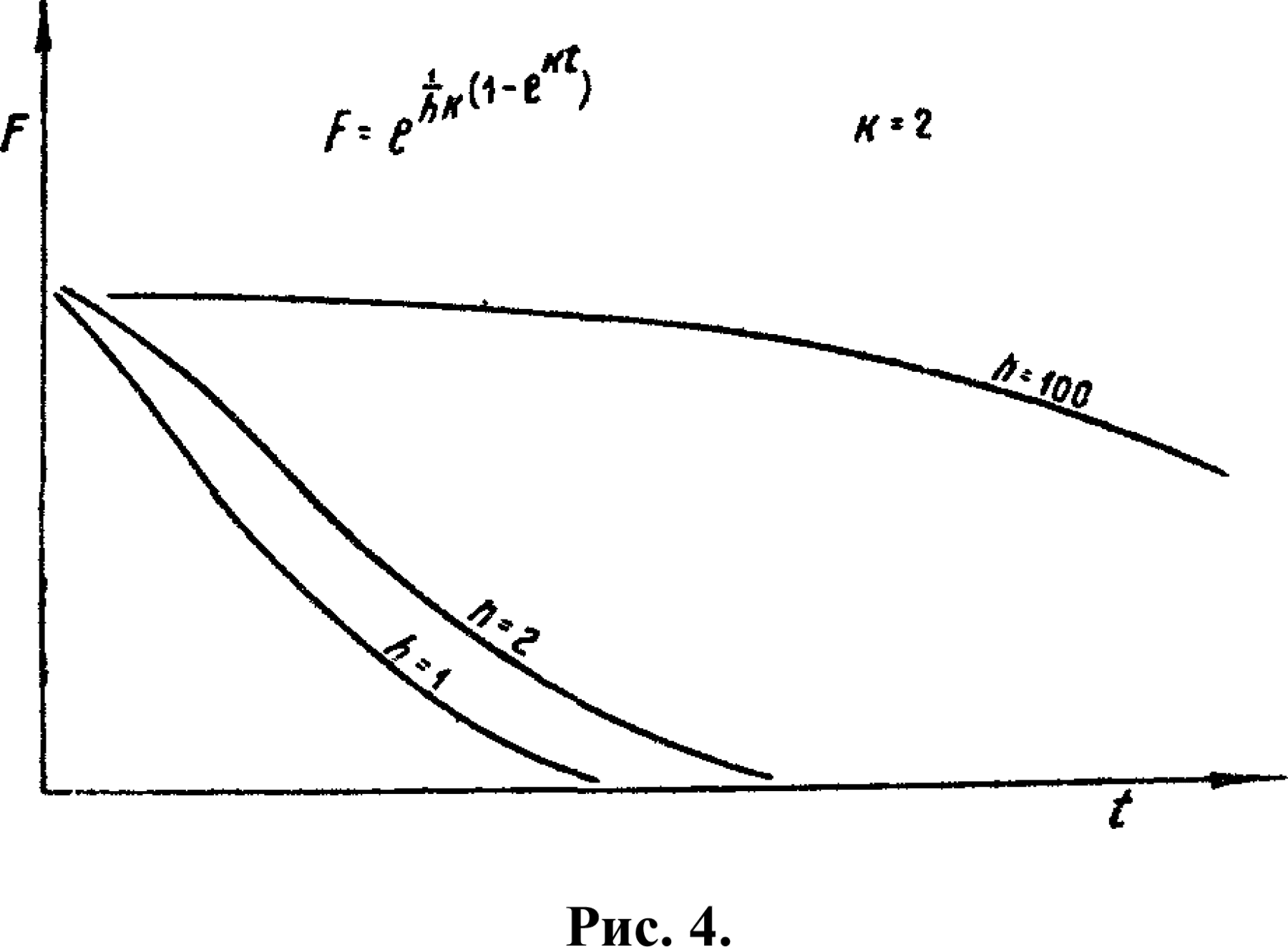
Thus, we see that the application of our laws of motion also leads to the result that the free energy of the living system, that is, the distance of the molecules of this system from the equilibrium position, must diminish after the attainment of the boundary of assimilation. Thus, the ageing of organisms is a necessity.
We see also that the manner in which this ageing proceeds in time, that is, the duration of life of the system at an identical initial value of (= the free energy of the living system after the attainment of the boundary of assimilation), depends on two constants: and . Namely, with an increase of the curve falls more quickly, while with an increase of —more slowly. This corresponds to the significance of the two constants. Indeed, whereas determines the speed of the processes of equalisation, independent of the vital processes, expresses the relation of the free energy of the system at each given moment to the intensity of the work of the structural forces (that is, to the intensity of metabolism) necessary for the preservation of the given level of free energy. Consequently, the higher is, the smaller the intensity of metabolism that will prove necessary for the maintenance of a certain degree of free energy at the given moment. Correspondingly, the potential of the system will also fall more slowly, that is, ageing will take place not so quickly; the duration of life will increase. On the other hand, it is clear that if increases, that is, if the processes of equalisation proceed more quickly, then with unchanged a more intensive work, a more intensive metabolism, is necessary in order to retard them; ageing will therefore take place more quickly, the duration of life will be shortened.
On the basis of our theory of living matter we already earlier arrived at the conclusion that the lowering of potential is an inevitable result of metabolism. To the same conclusion, as we see, our formulae also lead us. It therefore becomes clear too that the duration of life will depend on the intensity of metabolism, since the more intensive the latter, the more quickly is exhausted the free energy which living matter possesses in consequence of the deformed, non-equilibrium structure of its molecules.
On the basis of our theory of living substance we arrived at the conclusion that an inevitable result of metabolism is the fall of potential, and, as we have seen, the consideration of the adduced equations shows the same thing.
From our theory of living substance and of metabolism it follows, on the one hand, that upon the assimilation of a definite quantity of food substance a definite work must be developed at the cost of the free energy of the living substance; on the other hand, the energy of the food liberated thereby, in so far as it does not go to the building-up of new living substance, that is, in so far as there is no growth, must appear anew in the form of heat. Consequently, the free energy of the living system falls by a definite magnitude in relation to the quantity of heat formed by the acting mass of living substance.
Thus, we have:
where denotes the conversion of a quantity of heat, and a constant. The total quantity of calories that can be converted in the course of the whole of life at the cost of the free structural energy of the living system is determined by the expression:
that is,
We thus arrive at the proposition that the total quantity of calories that can be converted by the organism in the course of the whole of its life depends exclusively on the free energy of the egg-cell and is proportional to the latter. For the calculation of the quantity of calories that the organism can convert at the cost of its free energy from the moment of the attainment of the boundary of assimilation to the end of life, we obtain from this:
Since , where denotes the potential present at the attainment of the boundary of growth, then
where denotes the quantity of calories converted after the attainment of the boundary of growth. And since the mass remains here constant, then , and . This means that the quantity of calories that unit mass can convert after the attainment of the boundary of growth depends exclusively on the potential at the boundary of growth and is proportional to it. But , and especially for vertebrates, for example, the magnitude can be neglected in comparison with (the magnitude of the germ cell is negligible in comparison with the magnitude of the adult organism), and therefore (see Figs. 5, 6); we obtain:
This means that in the case where the mass of the egg-cell can be neglected in comparison with the mass of the adult organism, in this case the quantity of calories that can be converted by unit mass of living substance in the course of life after the attainment of the boundary of growth depends only on the potential of the egg-cell and is proportional to it.
M. Rubner[Author’s note:] M. Rubner, Das Problem der Lebensdauer und seine Beziehungen zu Wachstum und Ernährung, Verlag von R. Oldenburg, München und Berlin, 1908. showed that in various mammals of the most diverse magnitude the values of this ratio are approximately equal, whereas in man it lies higher.
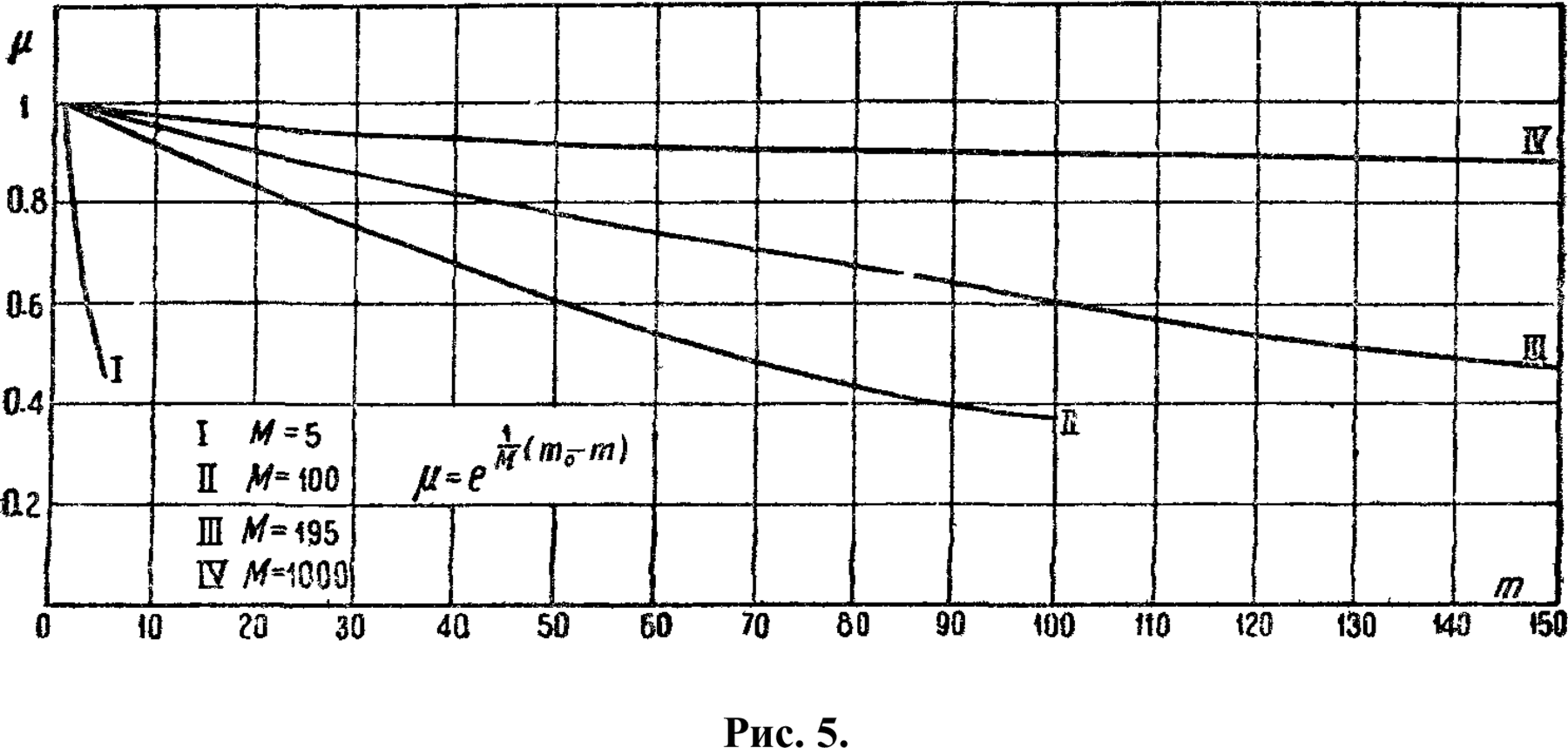
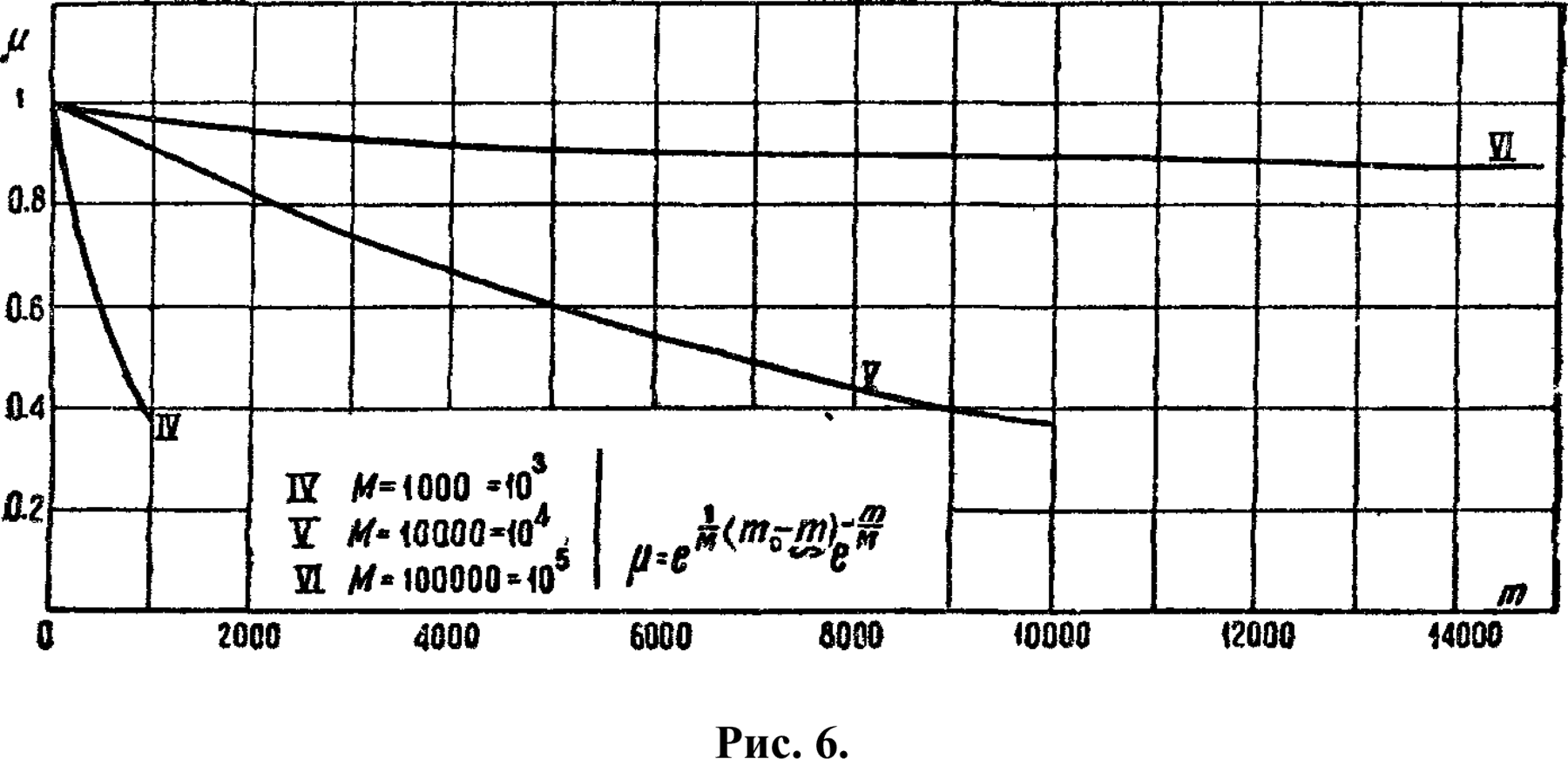
And from our proposition just deduced, taking into account that the magnitude can in the given case be neglected in comparison with , it follows that the regularity found by Rubner finds its explanation in the fact that the initial potential of the germ cells of the mammals investigated by Rubner is identical.
In one of my earlier investigations[Author’s note:] E. Bauer, Lebensdauer, Assimilationsgrenze, Rubner’sche Konstante und Evolution, Biol. Zbl., Bd. 51, 1931. I had occasion to point out that the magnitude increases in the course of evolution and remains approximately the same within each group. There too an attempt was made to generalise this proposition for another magnitude as well, in order to extend it also to the period of development and growth—namely, for the magnitude “the ratio of the work produced by the organism in the course of the whole of life to the free energy of the germ cell.”[Author’s note:] E. Bauer, loc. cit.
According to the considerations just developed, this magnitude is nothing other than , which was there named by us Rubner’s constant.
The numerical material adduced there, which was to illustrate in general outline the increase of this constant in groups appearing in later periods of evolution, concerned, however, the magnitude , the duration of life, including the period of growth, being taken into account. But if we wish to extend the regularity found by Rubner to the whole period of life, then we cannot speak simply of the total quantity of heat converted by unit mass, since in the course of growth the mass does indeed change.
We must in this case calculate the magnitude .
From our equation
we obtain, taking into account that
up to the boundary of growth
And since after the boundary of growth
then for the whole period of life, including the period of growth, we have
Thus, whereas after the attainment of the boundary of growth this magnitude depends on the potential and is proportional to it, for the whole period of life it is proportional not only to the initial potential but depends also on the magnitude determined by the boundary of growth.
The sense of all the adduced propositions amounts to the following. The magnitude , properly speaking, characterises by its definition the energy of activation of the molecules of food, the energy necessary for their assimilation. Since by all the data this energy of activation for different food substances, such as sugar, fat, protein, is different, the magnitude must depend on the dietary regime and is constant only with one and the same food.
If one neglects this dependence on the quality of the molecules of the food substances, or excludes it by the creation of a uniform dietary regime, then the magnitude can change in the course of evolution only in so far as the energy of activation necessary for assimilation changes. To direct measurement this magnitude does not as yet yield.
If we accept that the energy of activation necessary for assimilation of one and the same molecules of food is one and the same in different species, then in such a case the magnitude , earlier designated by us as Rubner’s constant, must remain constant. And then the magnitude , that is, the quantity of calories converted by the organism in the course of the whole of its life, can increase only as a result of the increase of the free energy of the germ cell. And in this case the magnitude , measurable by Rubner, of organisms in which the mass of the germ cell in relation to the mass of the adult organism is vanishingly small, can increase only with the growth of the magnitude , that is, of the potential of the germ cell. And since this magnitude does indeed exhibit a tendency to increase in the course of evolution, we shall retain the name of Rubner’s constant for this magnitude.
We shall return to it once more in the last chapter on evolution and in the chapter on adaptation and irritability.
Chapter 2: Reproduction
In the previous chapter we set forth the general regularities of metabolism and growth in living systems, and showed that these regularities are special forms of the manifestation of the general laws of motion of living matter. They represent the realisation of the possibilities contained in this form of motion, a realisation which leads to a more or less prolonged viability of living systems. But we saw that metabolism alone does not yet secure the permanent existence of living systems. The capacity for work of systems necessarily diminishes after the boundary of assimilation has been reached, living matter approaches the equilibrium state, and death must necessarily set in. In the present chapter we intend to show that within the form of motion of living matter, as it is expressed by our laws of motion, there lies the possibility of preserving the permanent viability of living matter even after the boundary of assimilation has been reached. Here we stand before a contradiction: on the one hand, it follows from our laws of motion that living systems cannot possess permanent viability, while on the other hand we assert that within these very same laws of motion there nonetheless lies the possibility of preserving such permanent viability. The resolution of this contradiction lies in the fact that, on the one hand, metabolism, in its realisation, does indeed prolong the life of the system, but at the same time necessarily leads to death; while on the other hand, metabolism is not the only possibility which the form of motion of living matter presents for prolonging the latter’s viability. In the introduction to this Part (p. 81) we have already indicated that living matter may, for a short while, exist as such even in the absence of metabolism, that is, the structural forces will perform work in it against equalisation, even without using the energy of molecules not belonging to the system. This, of course, can take place only in such a way that a part of the deformed molecules of living matter, being in non-equilibrium, increases the free energy, or the distance from the equilibrium position, of the remaining molecules at the expense of the energy of equalisation, that is, when a diminution of the living mass itself occurs, whereby simultaneously there may occur a raising of the potential of the remaining mass. Of course, if only this process takes place, then here too, after a definite time, death must set in, since it is accompanied by a diminution of the living mass. But it once again renders metabolism possible, metabolism being connected, as we saw, with a definite height of potential, and the potential falls as a result of metabolism. This process of raising the potential of one part of the living mass at the expense of another part of it is contained in our laws of motion as a particular case of them. It represents nothing other than the work of the structural forces of the system against the onset of equilibrium in it.
Thus we find the following connection between these processes: metabolism conditions the prolongation of the viability of living matter, but at the same time leads necessarily to death in consequence of the fall of the potential of the living mass which it occasions. The raising of the potential of one part of the living mass at the expense of another part of it likewise conditions the prolongation of the viability of living matter, but at the same time it too leads necessarily to death in consequence of the diminution of the living mass itself which it occasions. But since, as a result of the latter process, growth and metabolism again become possible, and since both processes (metabolism and the restoration of the potential) are contained as possibilities in the form of motion of living matter and are therefore particular cases of our general laws, then in the latter there is given the possibility of living matter preserving its viability permanently, that is, even after the boundary of growth has been reached. For this it is necessary that, after the boundary of growth has been reached, when the free energy of the living system begins to fall, but before the onset of death, the second process should regularly come into action — the process consisting in the alteration of one part of the living mass and in the raising of the free energy and potential of another part of it. Then metabolism and growth may once again take place, until the maximum of free energy (the boundary of growth) is once more reached, whereupon the potential again begins to diminish, the second process again sets in, and so on.
Thus the possibility of the permanent existence of living systems is inevitably bound up with cyclic changes.
This possibility, given in living matter itself, of preserving the viability of living systems even after the boundary of growth has been reached, is in fact realised in all the living beings known to us. But the form of this realisation in nature is not the one which appears the simplest from the theoretical point of view. The process of raising the potential of one part of living matter at the expense of another part of it — alternating cyclically with growth and ageing — is in nature usually bound up with an increase in the number of living systems and is realised in the most diverse forms in living beings which are, as it were, different from one another. But however diverse the processes of reproduction in different living beings may be, they are in every case founded upon one and the same phenomenon, namely upon the raising of the potential of one part of the living mass at the expense of the energy of another part of the mass of the living system.
Before passing on to a description of the various forms of reproduction as particular cases of this fundamental process [основной процесс — osnovnoi protsess], we must consider the process itself more attentively. As we have already said in the introductory Part, it is indeed to be seen as the fundamental process of living matter: for it is precisely this process that permits us to regard systems devoid of metabolism and wholly isolated from non-living matter as living, since, as a result of this process, equalisation is not permitted to occur without an alteration of the systemic conditions, but, on the contrary, work is performed against the onset of equilibrium at the expense of the energy of the structural forces of the system. Consequently, this process will always take place in those cases where metabolism is impossible. We then have before us a system whose non-equilibrium state is conditioned by the deformed non-equilibrium state of the molecules composing it — a system which is isolated from other molecules whose chemical energy it might use, by way of assimilation, for work. We have already said that in this case the living molecule would begin to approach the equilibrium state, which would unquestionably have set in without any alteration of the boundary conditions, since apart from this molecule there are no other work-capable structures, that is, no other work-capable parts of the system. The preservation, or restoration, of the non-equilibrium state by means of an influx of energy from without would signify a dynamic equilibrium with a source of work lying outside the system, that is, we should again not have a living system. The same would hold also in the case of a large number of such molecules of living matter, were there no possibility of interaction between them and were these molecules independent of one another. But if such interaction is possible, and the molecules are not independent of one another, then the energy liberated in the process of equalisation in one group of molecules may be used to increase the free energy, that is, the mean distance from the equilibrium position, or the potential, of other molecules, and we have before us a living system without metabolism. We shall not here enter into a consideration of what structural conditions are necessary for this interaction. In any case, we must here assume the presence of a very close connection of the molecules with one another. In the living systems now existing these conditions were undoubtedly fulfilled, for only such systems could indeed have been preserved; otherwise they would have had to die after a definite time even in the presence of metabolism. On the basis of what has been said above, we can represent this fundamental process, which serves as the basis for all the phenomena of reproduction, in the following manner.
If we assume that all the molecules at each given moment are in an identical state and, consequently, at an identical distance from the undeformed state of equilibrium, and if we denote the number of molecules in position I (before the accomplishment of the fundamental process) by , and in position II (after the accomplishment of the fundamental process) by , the molecular weight of the living molecules by , then, if the free energy is not diminished in the fundamental process, we have:
or, in graphical representation (Fig. 7).
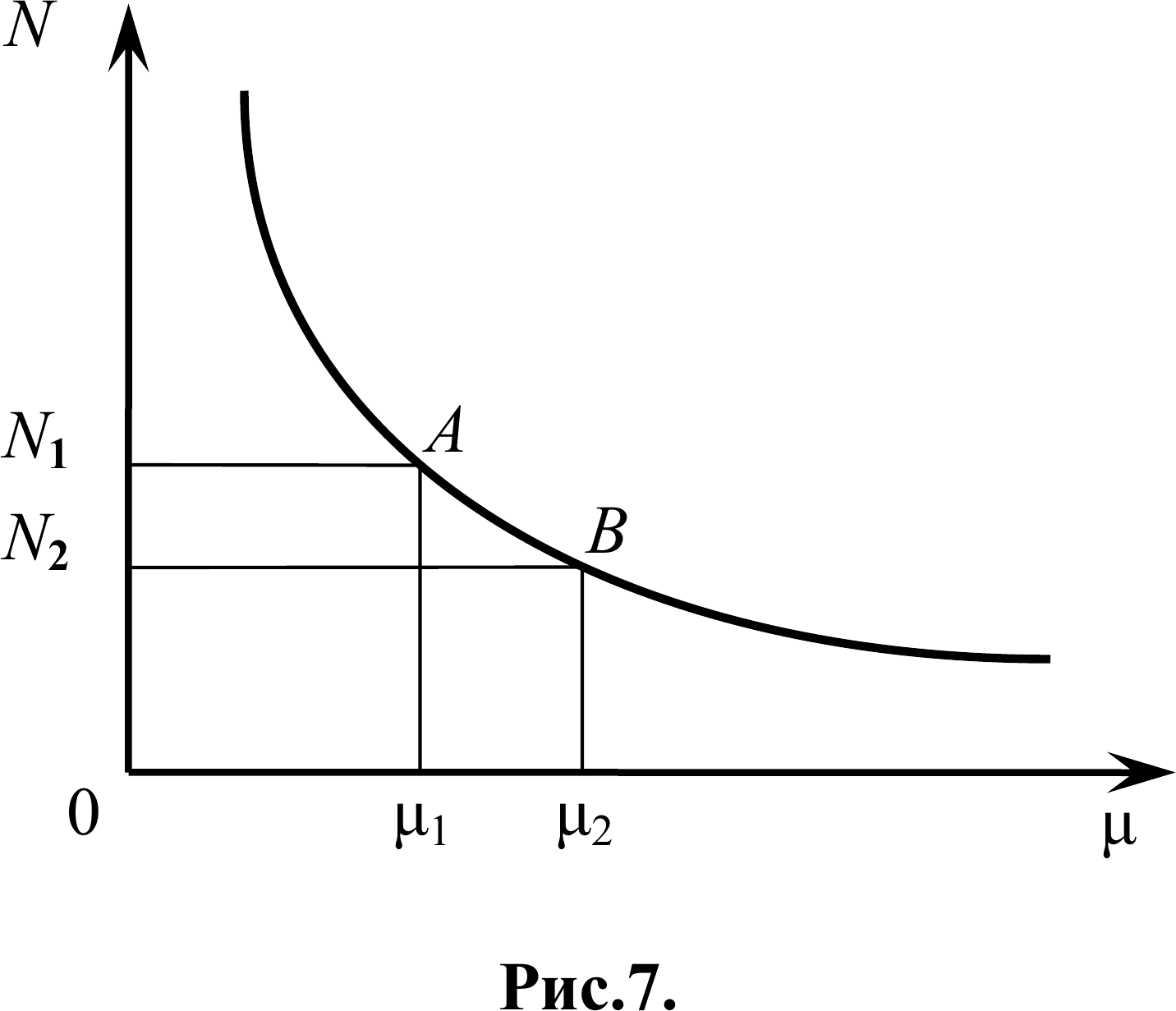
The areas of the two rectangles and must be equal. But in reality this is not the case, and the molecules in state I exhibit different distances from the equilibrium position. And for a given definite distribution of the molecules, their number in state I falling within the interval is equal to:
By we denote the number of molecules in state I as a function of . Thus, the total number of molecules will be:
while the entire free energy of the living system in state I will be
whence
In state II, that is, after the accomplishment of the fundamental process, the free energy will be expressed by
And since the molecular weight of the molecules does not change upon their excitation or deformation, and the free energy does not change in the fundamental process, then , and we have:
In what follows we operate with the mean values of the quantities at hand, so that, for example, for any state denotes the quantity which for state I we have denoted by .
We have already said that under natural conditions this process is encountered in the living beings now existing not in its theoretically simplest form, but always in conjunction with the process of reproduction. This becomes comprehensible if we recall that permanent viability is not fully secured even for that system in which the fundamental process proceeds periodically and thereby conditions this permanent viability: for a single chance fluctuation of the external conditions, exceeding the critical limits of physiological adaptability, is sufficient for our system to perish and to leave no traces whatever of its existence. We therefore cannot categorically assert that there did not arise and did not exist such systems as already possessed metabolism and periodically carried out the raising of the potential of one part of the system at the expense of the remaining parts, but did not yet reproduce.
In some organisms, however, we encounter this fundamental process, even at the present time, outside any connection with reproduction. Ordinarily this takes place under unfavourable external conditions, that is, when these conditions exceed the limits permitted by physiological adaptability, and when metabolism can no longer be carried out. This is the process of encystment, which we observe in many unicellular organisms and which is not connected with reproduction. But if in the organisms existing in nature the fundamental process is not encountered in its theoretically simplest form — that is, recurring periodically and thereby securing the permanent viability of the organism without any reproduction — then this, its simplest form, can be realised in many organisms by way of experiment. For this we must permit such an organism to metabolise up to the moment at which the fundamental process ought to set in. But we must prevent the onset of the process itself in its natural form, since it is, after all, connected with reproduction. Consequently, we must induce the fundamental process artificially before its natural onset. For this there exist two ways: we may, by halting metabolism, induce the equalisation of the deformed molecules of living substance, whereby, in consequence of the above-mentioned interaction, the energy liberated upon the equalisation of one part of the molecules will go to raise the potential of the rest; or else, by means of a local action, the destruction of the structure at a definite place, we induce equalisation in an insignificant part of the molecules; then, provided only that this structure is not specially adapted for some external work, the same process will set in, since the liberated energy of these molecules will raise the potential of the rest.
These two cases, however, differ sharply from one another in one respect. Whereas, when the possibility of metabolism is excluded — for example, by means of starvation — the fundamental process of raising the potential of one part at the expense of another must set in of necessity, since we are dealing with a living system, in the second case this is not obligatory. Namely, if by means of local destruction of the structure we liberate the energy of a part of the molecules, but do not exclude the possibility of metabolism, the fundamental process may also fail to set in. The non-equilibrium structure of the rest can be maintained by means of metabolism; the system may even continue to grow at the expense of the latter, for example, if it has not yet reached its boundary of assimilation. This difference becomes clear as soon as we recall our scheme of metabolism (p. 75). When metabolism is excluded as a result of absolute starvation, then we have deformed, active molecules, that is, molecules possessing residual valencies (), which must pass into the equilibrium state if they do not collide with the food molecules . But since this possibility is excluded in our case, then the fundamental process must set in, during which the molecules enter into interaction with one another approximately according to the following scheme.
In the second case, however, when by means of disturbing the structure we compel a part of the molecules to pass into the equilibrium state, but do not switch off metabolism in the rest, the fundamental process may also fail to set in. Here the food molecules will at the least compete with the molecules of living substance that are passing over to equilibrium and are deprived (in consequence of the disturbance of the structure) of metabolism, for affinity with the remaining molecules . But the organisms now existing in nature have preserved their permanent viability precisely thanks to the fact that they have realised the possibility, given by the laws of motion of living matter, of the metabolism necessary for their existence; therefore we must expect that in the living systems now existing the fundamental process will set in only in so far as preservation by means of metabolism is already impossible. Therefore, in such a competition, metabolism — that is, the binding of the free valencies of the molecules by the molecules — will prevail over the fundamental process — that is, the binding of such valencies by the molecules — for as long as this process leads to an increase of the free energy of the system as a whole, that is, until the boundary of growth has been reached. Anticipating what follows, we may already say here that it is precisely in this way that those facts are explained on the basis of which Hartmann and Jollos[Author’s note:] Hartmann, A. Allgemeine Biologie, S. 370, Jena, 1927. assumed that the cell is always capable of dividing, but is held back from division by growth. Since every process of reproduction, and consequently of division too, rests, as has been indicated, upon the fundamental process described by us, this theory of Jollos and Hartmann signifies nothing other than that the fundamental process can set in only after the boundary of growth has been reached. But if the possibility of metabolism, and consequently of assimilation, is excluded, then the fundamental process will set in of necessity and independently of the attainment of the boundary of growth, as a process characterising living matter in the absence of metabolism, on the ground of which we have designated it as the “fundamental process”.
As has been said, the fundamental process is in neither case obligatorily connected with the process of reproduction or division. This conjunction of the two processes is, as we have shown, a possibility which is contained in the form of motion of living matter and was realised in the course of history, this realisation being a necessary condition for the permanent viability of living matter and therefore having been realised in all the organisms now existing. By virtue of this, for the experimental, periodic induction of the fundamental process without reproduction — that is, for the experimental permanent preservation of the existence of living matter without reproduction — the necessary precondition is the possibility of inducing the fundamental process before its natural onset. The second precondition is, as we saw, the induction of this process upon the complete exclusion of metabolism, that is, by means of absolute starvation; whereas, if we induced it by means of the direct destruction of the structure of one part, then to this is added a new condition, namely that we must induce the fundamental process after the boundary of growth has been reached.
We know that we do indeed succeed in preserving, in some organisms, permanent viability without reproduction, by means of periodic starvation or periodic local disturbances of the structure. Thus Goetsch,[Author’s note:] Goetsch, W., Biolog. Zbl., Bd. 41 and 42, 1921 and 1922. as is known, was able, by means of periodic starvation, to maintain life permanently in hydras, while Hartmann[Author’s note:] Hartmann, Biol. Zbl., Bd. 42, 1922; Naturwissensch., H. 19, 1926; Zool. Zbl., Abt. f. Zool. und Physiologie der Tiere, Bd. 45, 1928. succeeded in achieving this with Stenostomum by means of repeated amputations, these organisms not reproducing. Hartmann achieved the same thing also with the unicellular Stentor and Amoeba proteus, likewise by means of amputations. The experiments of Cheifec[Author’s note:] Cheifec, Arch. f. Protistenkunde, Bd. 70, H. 1, 1930. speak in favour of the possibility of attaining the same results in unicellular organisms also by means of starvation, since he succeeded in making a culture live without reproduction by means of appropriate dosing of nutrition.
A direct experimental proof of our theory concerning the mechanism of this phenomenon is given by the experiments set up in my laboratory and under my direction by A. M. Granovskaya.[Author’s note:] E. S. Bauer and A. M. Granovskaya, Biol. zh., t. III, no. 3 and 4, 1934. The object served was a clone of the infusorian Oxytricha, which was kept in a hay infusion. First of all, we succeeded in showing that to any operation — be it the cutting off of a part or only an incision, that is, to any local disturbance of the structure — Oxytricha responds by a reconstruction of the nuclear apparatus, which is analogous to the physiological processes of “rejuvenation” in endomixis, or parthenogenesis. Consequently, these processes may serve as the morphological expression of the fundamental process having set in. If the above representations concerning the role of this process and its relation to metabolism are correct, then such a process of reconstruction, as a reaction to local injury, ought not to set in at any stage whatever of the life of the cell. It ought not to set in in young cells which have not yet reached their boundary of assimilation, and it ought always to set in in old cells which have already reached this boundary or have passed beyond it, but have not yet passed over to normal reproduction. Our object, under the given conditions, divided at room temperature twice a day, that is, it possessed a division period equal to 10–12 hours. We therefore carried out a large number of operations (incision or excision of a small piece) on individuals 6–7 hours after
division, assuming that, with the stated division period, the boundary of assimilation had by that time already been reached. Of two sister cells, one was operated upon, while the other served as a control; after a definite time following the operation, both cells were simultaneously fixed and stained. The same experiments were then carried out with other individuals immediately, or half an hour after, they had divided, proceeding from the assumption that these cells had certainly not yet reached their boundary of assimilation. In all the numerous experiments, cells at an age of 6–7 hours always reacted by a reconstruction of the nuclear apparatus, which remained on the whole everywhere the same. Younger cells, which had not yet reached their boundary of assimilation, never reacted by a reconstruction of the nucleus; the nuclear apparatus remained in just as complete a state of rest as in the sister cell which served as a control.
Above we indicated that upon disturbance of the structure, metabolism continues up to the attainment of the boundary of growth, and the fundamental process does not set in, in consequence of which assimilation predominates; whereas after this boundary has been reached, metabolism does not take place, and there sets in a raising of the potential of the molecules at the expense of the dissimilation of the disturbed molecules torn out of their connection. Therefore one should expect differences also in the reaction of metabolism to operative action. On the basis of these considerations we set up the same experiments, but with a determination of respiration (oxygen consumption) in the operated and non-operated (control sister) objects. The determination of respiration was carried out by the method of Kalmus. These experiments showed that indeed in operated adult organisms the respiration immediately after the operation, and an hour after it, was higher than in the control individuals, whereas in young organisms the operation always induced a lowering of the intensity of respiration. Upon the determination of the respiration of operated adult individuals 3 hours after the operation, however, we found a lowering of the intensity of respiration, that is, the metabolism in these individuals was already the same as in young organisms, in which assimilation, having begun anew, predominates over dissimilation.
Thus these experiments give a direct experimental proof of our theoretical propositions concerning the relations between metabolism and the fundamental process. From them it directly follows that so-called experimental individual immortality can be realised only in the case where the corresponding lesion — the operation — is inflicted after the boundary of assimilation has been reached. If, however, we operate before this boundary is reached, then we must expect the opposite result, namely a more rapid ageing, that is, a shortening of the life cycle, since in such a case the operation will induce an intensification of assimilation, and consequently also a more rapid fall of the potential in the protein molecules. That this is indeed so was shown by our experiments.[Author’s note:] Loc. cit.
We see that in individuals which have not grown to full size, repeated operations constantly lead to a more rapid division and to death, whereas in adults they hinder division and considerably prolong the preservation of living matter without reproduction.
But we have here no right whatever to speak of the immortality of one and the same individual. This is evident from the fact that immortality here is in essence conditioned by the very same process, the very same mechanism, as reproduction, that is, the arising of new individuals, and that such immortality can be induced only in so far as one induces this process. We are here dealing with the cyclic induction of a complete reconstruction and a raising of the potential at the expense of the system’s own structural energy upon the temporary switching-off of metabolism, that is, with the formation of one, and not of two or several, individuals, as would normally be the case in the absence of experimental intervention. The reconstruction of the individual by means of the fundamental process, upon which all reproduction rests, is induced experimentally, whereby only the phenomena of reproduction connected with this process are disturbed.
Thus we have deduced the relations between metabolism and the fundamental process in its connection with reproduction from our general laws of motion, without having recourse to any special, restrictive assumptions. We must therefore accept that these relations, although in different forms, are encountered in all living beings, that is, in multicellular organisms as well as in unicellular ones. We have already said that, by means of periodic starvation or periodic operations, it is possible to induce so-called individual immortality also in multicellular organisms. But these are exactly the same actions as those which led (and, on the basis of our theoretical considerations, ought to have led) to similar results in unicellular organisms. The assumption of fundamentally different mechanisms and relations for this case would therefore be, at the very least, a great strain. But of course the forms in which this process manifests itself in unicellular and multicellular organisms, or in plants and animals, are so different that it is often difficult to discern, within this diversity, the historically changing and developing manifestation of one and the same regularity.
It is therefore comprehensible that Weismann, proceeding from the observations and the level of science of that time, considered that between unicellular and multicellular organisms there exist fundamental differences with respect to reproduction and immortality. In view of the fact that he could note only the various manifestations of the processes of reproduction, but could not know the general regularities upon which they are founded, he was able to arrive at a division of living matter into two parts — into the soma and the germ plasm [зародышевая плазма — zarodyshevaia plazma], of which the first is mortal, and the second immortal. If we cast aside our representations concerning the relations between the fundamental process and metabolism, and also concerning the fundamental process as the most essential manifestation of the laws of motion of living matter, then on the basis of direct observations we shall necessarily arrive at such a division of living matter into two parts. For directly we always observe that all multicellular organisms reproduce by means of germ cells, whereby the cells of their body die, and only in unicellular organisms, in which the somatic cell so to speak coincides with the germ cell, there sets in not death, but reproduction (division of cells). Above we adduced experimental proofs that so-called “immortality” is connected with the fundamental process, that is, with the dying-off of one part of the living mass and with the reconstruction of another part of it, leading to a raising of the potential. The experiments of Hartmann and Goetsch on multicellular organisms show that here too we are dealing with the very same process. Consequently, reproduction is based, in multicellular organisms as well as in unicellular ones, upon one and the same fundamental process. But then the necessity of dividing living matter into a mortal soma and an immortal germ plasm falls away. The continuous new formation of germ cells from generation to generation in multicellular organisms takes place in consequence of the process of the dying-off of the cells of the body at the expense of the structural energy of the latter, just as the reproduction of unicellular organisms takes place at the expense of the structural energy of those parts of the cell which thereby inevitably die. This connection between the germ cells and the somatic cells follows directly from our proposition, according to which every process of reproduction rests upon the fundamental process; this proposition, in turn, we obtained by way of the application of our general laws of motion of living matter. With their help we can not only explain the reproduction of unicellular and multicellular organisms without having recourse to a division of living matter into two parts, but can also understand a whole series of other facts not explicable from the standpoint of Weismann’s positions. To these belong, in the first place, the above-adduced experiments on so-called individual immortality in unicellular and multicellular organisms, and, in the second place, a whole series of facts showing that, in the animal kingdom as well as in the plant kingdom, there exists a close dependence between the duration of life and the dying of the body, on the one hand, and reproduction, or the formation of mature germ cells, on the other. Thus, for example, in worms the formation of germ cells is connected with death; in fishes there is observed the disappearance of great masses of the body upon the spawning of roe, and so on. Among plants, let us mention certain species of palms, which grow for many years without bearing flowers, and then, having flowered, die. By way of experiment it is also possible to show the presence of a connection between the formation of germ cells and dying in plants. Thus, for example, it has proved possible experimentally to prolong life in species of Pelargonium, Draba, Veronica, Reseda, Lobelia, Conium maculatum, by artificially delaying flowering by means of cutting off the branches that form buds. All these facts find their direct explanation on the basis of our theory, according to which the germ cells, with their maximal initial potential, are formed at the expense of structural energy, that is, at the expense of the dying — in other words, the dissimilation — of the tissues of the body. From the standpoint of Weismann’s theory, however, these facts cannot be explained.
But if reproduction in unicellular and multicellular organisms is indeed founded upon one and the same fundamental process, then it manifests itself in different species, as we have already said, in the most diverse manner. This depends upon the manner in which the above-described connection between the fundamental process and metabolism is realised in dependence upon the peculiarities of structure in different species. Therefore we shall, proceeding from our general laws of motion, first investigate the theoretical possibilities at hand, and then look at which of them are realised in which cases. We are able to do this already quite easily and graphically. Let us draw for this purpose a -diagram (Fig. 8), depicting the dependence of the potential on the mass for metabolism and for the fundamental process. The family of curves depicting metabolism is, as is known, represented by the type with the parameter . They show the fall of the potential upon an increase of mass. The family of curves depicting the fundamental process belongs to the type of equilateral hyperbolae, since in the fundamental process the entire structural energy of one part of the system is used for the raising of the potential of another part of it. That the curves of metabolism are less steep follows already from the fact that in assimilation the structural energy of the system itself, necessary for the rebuilding of non-living mass — that is, of mass not belonging to the system — is used, whereas in the fundamental process the remaining mass, already belonging to the system, is rebuilt at the expense of this structural energy.
There exists a certain analogy between this -diagram and the -diagram of gases. The curves of metabolism correspond in this case to the isotherms (which, however, in gases are represented by hyperbolae), while the curves of the fundamental process correspond to the somewhat steeper adiabats; the potential corresponds to the pressure, and the mass to the volume. As there, along an isotherm, an exchange of heat takes place upon a change of volume, so here, along a curve of metabolism, changes of mass take place upon a chemical exchange of energy and substances. Corresponding, in turn, to the absence of an exchange of heat and indeed of all energy upon changes of volume along an adiabat, we here have the absence of metabolism upon changes of mass along a curve of the fundamental process. But here the analogy comes to an end. Indeed, proceeding from the proposition that all changes take place only through equilibrium states, we draw, for gases, the conclusion that all processes are reversible and that we can move in both directions — both along the isotherms and along the adiabats. For living systems, however, on the basis of our proposition that we are here dealing only with non-equilibrium states, movement is possible only in one direction, namely — as the arrows on the figure show — we can, according to our laws of motion, move along the curves of metabolism only to the right, and along the curves of the fundamental process only to the left. A cyclic process along these curves — that is, by means of a combination of metabolism and the fundamental process — is therefore impossible.
Another restriction lies in the fact that, according to the considerations expressed in the chapter on metabolism, the diagram remains real only up to the boundary , that is, along the curves of metabolism we can proceed only as far as the boundary of growth. In accordance with the equations derived in the chapter on metabolism, according to which growth beyond the boundary of growth would already lead to a diminution of the free energy, the line is distinguished by the fact that up to it every curve of metabolism intersects ever new curves of the fundamental process, whose energy increases ever more, while after it the curve of metabolism intersects the curves of the fundamental process once again, but now with a lower energy. Consequently, to each curve of metabolism there belongs only one hyperbola (curve of the fundamental process), with which it makes contact at ; all those lying lower it intersects once again after this line. This follows from the following calculation.
The equation of the curve of metabolism has the form:
The equation of the curve of the fundamental process has the form:
The equation of the points common to both curves has the form:
The question thus reduces to the following: in how many points is the curve of the function intersected by the straight line ? Either in none, or in two, or the straight line will be tangent to the curve in one point. At the point of tangency
Hence there follows for our functions
where denotes the abscissa of the point of tangency.
Let us determine and . From the cited equations there follows:
Consequently, to the curve of metabolism
there belongs the tangent hyperbola
States to the right of the boundary line are biologically not real, but states along this boundary line are real; these are the states through which the system passes after the attainment of the boundary of assimilation at constant mass, that is, in stationary metabolism. As we have shown, the maintenance of this state too is inevitably accompanied by a fall of the potential in the course of time. Consequently, the boundary line too can be traversed only in one direction, namely — as the arrow on the figure shows — from top to bottom.
But if in this way a cyclic process is impossible along the curves of metabolism and of the fundamental process, then it can nevertheless be realised along the curve of metabolism, the boundary line, and the curve of the fundamental process — for example, as is depicted in Fig. 8, that is, in the direction .
This simple “cyclic” process without reproduction is realised experimentally in the above-adduced experiments on so-called individual immortality. The necessary condition for this is, as is at once evident from our -diagram, the inclusion of the process of the fall of the potential without growth between the increase of mass and the fundamental process. The latter ordinarily sets in only after the attainment of the boundary of assimilation, but experimentally — for example, by means of starvation — it can be induced also earlier.
Theoretically we can represent to ourselves the following cases of combinations of the fundamental process and metabolism without reproduction: a) a raising of the potential at constant mass, approximately corresponding to the following combination of metabolism with the fundamental process: . But this process inevitably leads to ever smaller oscillations, as is represented in Fig. 9, and consequently cannot take place without limit; b) an increase of mass at constant potential, corresponding to the following combination of the two processes (Fig. 10). This process too is limited, both in consequence of the diminishing oscillations and in consequence of the attainment of the boundary line of the boundary of growth, provided only that during the fundamental process there does not take place so deep a rebuilding that, in consequence of a change of , the boundary of growth itself also changes.
All these three cases, not accompanied by reproduction, have no significance for evolution and, moreover, are not encountered in nature. But theoretically they are possible, and therefore it must be supposed that they can be realised by way of experiment on suitable objects and under suitable conditions. As regards the first case of a cyclic process, such experiments, as we saw, already exist, whereby in our experiments with Granovskaya the cyclic process set in precisely under those conditions which the theory requires. As regards the other cases, there are as yet no experimental data concerning them, but in part — and in combination with reproduction — all the cases are realised in nature. The first we encounter in the formations of cysts, serving for rejuvenation under unfavourable conditions; the rest, the meaning of which in essence reduces to the same thing — namely to the inclusion of the fundamental process before reproduction and before the attainment of the boundary of growth — we find in nature under the guise of metamorphosis. The role of metamorphosis in evolution is very diverse. In the first place, the inclusion of the fundamental process gives the organism a possibility, very advantageous in the ecological respect, of rebuilding the organism; in the second place, this rebuilding may push back the boundary of growth by means of a change of the coefficient , that is, of the work necessary for the assimilation of a unit of mass, which may signify a prolongation of individual life or an increase of external capacity for work; finally, the inclusion of the fundamental process may, in consequence of the raising of the potential, lead to the formation of a greater quantity of germ cells, that is, a more numerous progeny.
In favour of the fact that in metamorphosis, and in the histolysis taking place during it, we are indeed dealing with the fundamental process, there speaks also the fact that, for example, according to Krogh[Author’s note:] Krogh, Respiratory exchange in man and animals, 1916. this histolysis is accompanied by a raising of respiration, just as was the case in our experiments on the operated, reconstructing adult individuals of Oxytricha.
The rise of the intensity of respiration in the last days before emergence from the pupa is to be attributed to an intensification of movement, but for the first period this cause can be considered excluded. Some authors explain this heightened intensity of respiration in the first period by the processes accompanying histolysis. Krogh, on the contrary, considers it possible that here a diminution of mass takes place, which is what causes the lowering of respiration. But against this objection of Krogh’s there speak our experiments on Oxytricha, where in operated individuals, at times constituting no more than a third of the entire organism, respiration was nonetheless heightened several times over. We therefore consider Krogh’s supposition improbable also with respect to metamorphosis. Thus in our -diagram metamorphosis will be represented by the scheme (Fig. 11).
The different course of the curves and must indicate that, in consequence of the deep rebuilding in metamorphosis, there may also occur a change of the parameter , that is, of the boundary of growth. Thereby the lattice of curves is, as it were, deformed.
Thus we see that all the theoretical possibilities of combinations of metabolism with the fundamental process without reproduction, as they are represented on our -diagram, were indeed realised in nature in the course of evolution, in so far as they had evolutionary significance. As regards the processes of reproduction, we shall likewise consider them on the basis of our diagram. The theoretically simplest case is, of course, that in which the fundamental process sets in immediately after the attainment of the boundary of assimilation and leads to a doubling of the mass with the initial potential. This is the minimally necessary condition for any unlimited reproduction, for if the fundamental process did not lead at the least to the restoration of the initial potential, then we should obtain a series of generations with an ever-diminishing potential, which would inevitably lead to the extinction of the given species. And if at the least a doubling of the initial mass were not attained, then this would lead to a constant diminution of the magnitude from generation to generation, and consequently to a diminution of all the free energy, and as a result likewise to extinction. The elementary, uncomplicated process of reproduction is therefore represented, in our diagram (Fig. 12).
From our equations it follows that the potential falls with growth; in consequence of this the mass must increase by more than twofold in order that the free energy should double. And since, for the attainment of a double mass with the same potential, the free energy must double, it is clear that the process of reproduction is possible only in the case where the boundary of growth exceeds the initial mass by more than twofold. This is at once evident from our diagram: in order, going from the initial point along the curves of metabolism and of the fundamental process, to reach the point , we are compelled, in the course of the curve of metabolism, to pass over to the abscissa . This elementary process of reproduction is realised in all unicellular organisms reproducing by means of simple division. We thus arrive at a very general proposition, according to which in every unicellular organism reproducing by means of division the condition must likewise be realised, without which the culture perishes. That this condition is fulfilled in multicellular organisms and in the majority of unicellular ones, we can, it would seem, satisfy ourselves by direct observation, even without making exact measurements. In other protists, however — for example, in bacteria — this question stands in need of more detailed investigation, the more so as concerning them there prevails the conviction that the rods grow to a double length without increasing in thickness, and then divide into two equal rods possessing the initial magnitude.
Calculating approximately the minimal excess of the magnitude as compared with , we obtain the following: to the curve of metabolism
there belongs the hyperbola
satisfying the conditions:
- the hyperbola intersects the curve at ,
Consequently
This condition signifiesThe 1935 original prints the numerator as ; from condition (2) and the preceding equation it should read — evidently a misprint, which the next equation tacitly corrects (using ). Reproduced here as printed.
Whence there follows:
And since and denotes the ratio of the maximally increased mass to the initial mass, then
which gives the approximate value
that is,
At first sight it may seem that this condition is not fulfilled, for example, in bacteria. If this is indeed so, then according to our theory with each generation there must take place a gradual diminution of the free energy of the individuals, and, in the end, the culture must perish. We know that cultures of bacteria do indeed exhibit phenomena of ageing; their virulence diminishes. In so far as exact investigations exist, they show that the lowering of the free energy manifests itself also in a lowering of the capacity to assimilate. Thus, Kunstmann (on Aspergillus niger), Noak (on Thermoascus aurantiacus) and Tausson (on Aspergillus flavus)[Author’s note:] Cited after Tausson, Mikrobiologiya, t. II, 1933. showed that from the same quantity of nutritive substances an old culture forms less cellular substance than a young one, that is, if denotes the weight of the dry substance of the organism formed, and the weight of the nutritive substances consumed, then the ratio diminishes in proportion to the ageing of the cultures. But if cultures do age, then this takes place only under laboratory conditions; or — if ageing takes place also in nature — they must somehow rejuvenate themselves anew, that is, they must, after a definite number of divisions, again form individuals possessing the free energy — that is, the mass and potential — of the initial form, without which the given species must perish. But this means that there must nonetheless arise individuals whose mass is at the least 4.5 times greater than the initial one, that is, the magnitude of the adult cells in a young or rejuvenated culture must be at the least 4.5 times greater than the magnitude of the cells in an old culture no longer capable of growing and reproducing. Bayne-Jones, Stanhope and Edward F. Adolph[Author’s note:] Journal of comparative and cellular Physiology, no. 2, 1932. found precisely this magnitude, investigating Megabacterium by means of the microcinematograph. They found that the volume of the adult cells falls progressively from the beginning of cultivation and, towards the end of the period of reproduction, attains only 1/5 of the magnitude of the initial maximum.
Ordinarily, bacterial cultures which have aged again acquire their capacity to grow and their virulence upon re-inoculation. One should therefore think that the lowering of the boundary of assimilation is induced precisely by the conditions of the culture, since no longer all the cells out of the number living in one culture can attain their natural boundary of assimilation. Many of them prove to be damaged, in consequence of insufficiency of nutrition, accumulation of products of metabolism, and so forth, even before reaching this boundary, and thereby the fundamental process leading to division is induced in them. In a culture, consequently, we have before us not the simple process of division in its pure form, as it is depicted in Fig. 12, leading to the formation of two cells equal in value to the initial one, with identical potential and mass, but we have, alongside it, the appearance of the fundamental process leading to division before the attainment of the boundary of assimilation. This is represented in our diagram in Fig. 13, that is, besides the normal process of division , there take place in the culture “premature” divisions . From the diagram it is at once evident that such divisions can lead to the initial potential only in the case where the daughter cells are smaller than the maternal one, since the abscissa is necessarily smaller than ; if, however, the division leads to the formation of daughter cells of mass identical with that of the maternal one, then their potential, and consequently the boundary of growth, is lower, since the ordinate is smaller than . Consequently, cells dividing “prematurely” must produce an ever smaller or ever more poorly assimilating progeny. By this, and by this alone, can the ageing of cultures be explained. For if all the cells in a culture also divided according to the normal scheme , then the lowering of the boundary of assimilation and the diminution of the magnitude of the cell could not take place. In the literature, it is true, there are not encountered exact, systematic investigations of the individual variability of the magnitude of cells in cultures of unicellular organisms and of their changes in connection with the ageing of the culture. But in our own, not yet completed, investigations on lactic-acid bacteria we have established a considerable variability of the magnitude of the cells in one and the same culture. The same follows also from the mentioned works of Bayne-Jones and others.[Author’s note:] Loc. cit.
These authors adduce, for example, two individual growth curves from one and the same culture, from the moment of one division to the moment of the next. From these curves it is evident that whereas one bacterium grows to approximately double the size and then divides, another, before dividing, attains more than threefold the magnitude.
But if, on the one hand, the ageing of cultures is indeed a very widespread regularity, then, on the other hand, we know that by means of appropriate methods cultures can be kept alive for an unlimitedly long time, and that, besides, in nature various species of unicellular organisms can ordinarily avoid extinction. The presence of cells which attain their normal boundary of growth and are therefore able to create, as a result of the fundamental process leading to division, two new cells with the initial potential and the initial mass, is, consequently, a necessary condition for the unlimited viability of cultures of unicellular organisms, since these cultures live unlimitedly precisely at the expense of such cells. But then the question arises: by what is the constant presence of such cells with the initial potential secured? Since in freely living unicellular organisms reproducing by means of division we regard each cell as an independent living system, and therefore can assume the existence of the fundamental process only within a single cell, it is entirely evident that cells with the initial potential and mass — that is, with the normal boundary of growth — can arise either by way of division from cells of the same kind, forming identical products of division, or else we must assume the existence of a division leading to the formation of non-identical products of division. In the latter case, from a cell with premature division, or from a cell arising from it with a lowered boundary of assimilation, there may arise one cell with the initial potential and mass. According to our scheme this is possible in the case where the cell, being in stage , upon development corresponding to the curve of the basal metabolism , divides according to the formula:
An unequal division in the sense of the formation of daughter cells with identical mass, but with different potential, would mean that during division a regrouping of the material in the cell had taken place, or else that in the latter, from the very beginning, there had been differences in the structure of the separate regions. Finally, there exists yet a third theoretical possibility: in a cell with a lowered boundary of assimilation (but necessarily lying above ) there sets in, after the attainment of this boundary, the fundamental process, but without division, that is, with a raising of the potential above the initial level for the given cell. This case is analogous to the rebuilding induced in our experiments by means of traumatisation, and is encountered in nature in ageing cultures of infusoria in the form of endomixis.
Thus we can represent all three possibilities of the unlimited reproduction and prolongation of life in cultures of unicellular organisms in the following manner:
- On the assumption of equal division (that is, the formation of identical products of division).
- On the assumption of unequal division
(that is, the formation of non-identical products of division).
- In the case of endomixis.
In these figures the black circles denote cells with the initial potential and mass, the shaded ones — cells with a lowered potential, or smaller mass, the white ones — cells with a still more lowered boundary of growth, and, finally, the circles with a cross — cells whose boundary of growth lies below and which must therefore perish.

Which cells in a colony of freely living unicellular organisms divide normally and form products of division with the initial potential and mass, and which, in consequence of insufficiency of nutrition or other damage, divide prematurely and form products of division with a smaller free energy — this depends upon chance, for indeed all cells with the initial energy are equal in value. The number of damaged cells and their percentage ratio to the cells with the initial energy will therefore likewise be determined by statistical regularities. It is self-evident that with an increase of the density of the culture, or with a diminution of the quantity of living and non-living nutritive substances at the disposal of the organisms — perhaps also in connection with an increase of the products of metabolism that may exert a harmful action — the percentage of full-value cells will diminish. From this it follows, on the one hand, that under unfavourable conditions unequal division, or endomixis, will set in earlier, and, on the other hand, that so long as the density of the culture is small — that is, immediately after its sowing — the first divisions will in all probability be normal and will lead to the formation of daughter cells with the initial energy.
This chance circumstance, founded upon the equality of value of cells with identical free energy in cultures of unicellular organisms, passes over into a regularity in colonies, and still more so in multicellular organisms. In multicellular organisms this regularity consists in essence in the fact that the various cells in successive cell generations are already “not equal in rights”. Multicellular organisms no longer represent a culture of independent living beings — cells — that influence one another, but are themselves living systems, for which therefore all the laws of living systems retain their significance. It is precisely by virtue of this unity of the whole system that the cells composing the multicellular organism, or the successive cell generations in the development of the latter, are not equal in value. These cells have each their own destiny: they acquire, so to speak, their individuality at the price of the loss of independence. Thus, in the limiting case, this regularity consists in the fact that in each cell generation, beginning with the stage of two blastomeres, each cell can receive an individual designation or number, whereby, on the basis of this designation or number, the number and kind of cells forming from it in the further cell generations can be indicated. To what extent this regularity is realised in different organisms, and by what such a determination of cells is conditioned — with this is occupied the mechanics of development founded by W. Roux.
This question interests us here only in so far as we must investigate to what extent the regularities of reproduction set forth by us are realised in multicellular organisms. But before passing on to multicellular organisms, we must consider some intermediate cases. Let us dwell first on the theoretically simplest case, when as a result of division there arise not two systems with the initial free energy, but several such systems. For this there is needed only a potential of the system so high that it attains the boundary of growth when its free energy exceeds the free energy of the initial form by several times. In our -diagram this case is represented in the following manner (see Fig. 17).
This case is realised in nature in so-called multiple division, encountered in Foraminifera, Radiolaria, Sporozoa and many algae. Since in these cases the diminution of mass becomes noticeable morphologically only after the division of the nucleus, it is to be supposed that the fundamental process here indeed begins after the attainment of the boundary of growth of the multinuclear formation, and that by virtue of this each of the products of division acquires its initial potential only subsequently, already after the disintegration. Another intermediate case is the formation of colonies, where only the number of divisions is determined, while the divisions themselves proceed according to the fundamental scheme and form products equal in value with the initial potential and mass. Finally, we know cases where not only is the whole number of divisions determined, but, after a definite number of them, in a definite quantity of the products of division there set in premature divisions, leading to the formation of cells with a lowered potential, that is, a lowered boundary of growth. This takes place, for example, in Pleodorina, where of 32 cells 28 are full-value, while 4 possess a lowered boundary of assimilation and have already lost the capacity for further division. Here we must assume one of two things: either the divisions take place according to scheme 1, whereby products of division equal in value are formed, but at the stage of 8 cells one of them divides prematurely, before the attainment of the boundary of growth; on the assumption of the formation only of products of division equal in value, this premature division could not set in either earlier or later, since in none of the other cases can the ratio 28 : 4 appear. The other possibility lies in the assumption of unequal division (the formation of products of division not equal in value), approximately according to our scheme , by which the difference in the magnitude of the cells may likewise be explained. In such a case this unequal division must set in already at the stage of 4 cells, one of which, upon its division, forms unequal cells. Thus there arise 7 full-value cells and one with a lowered boundary of assimilation. Then, by means of two equal divisions, from the first there are formed 28 full-value cells, and from the last — 4 somatic cells incapable of dividing. Here the question arises, by what is it conditioned that in the first case one of the 8 cells does not attain its natural boundary of growth and divides prematurely, while in the second, that one of the 4 cells divides unequally? And further, is it predetermined in advance which of the 8, resp. of the 4 cells, divides prematurely, resp. unequally? From our theory it follows that in the first case all 8 cells must be equal in value; therefore the premature division of one of them can be induced only by the fact that under the given conditions not all the cells are equally provided with nutrition. Thus, that one of the 8 cells which is to divide unequally can be determined only by its position, and not by some hereditary property. In the second case our theory allows two possibilities: either all 4 cells are still equal in value and one of them divides unequally by virtue of the same causes which induce also the premature division of one of the 8, or else one of the 4 cells is already not equal in value to the remaining three and possesses a lowered boundary of growth, that is, is able, even on condition of attaining this boundary, to give only one full-value product of division, while the second will necessarily possess a smaller mass, or a lowered potential. But from this it follows that already the first division must have been unequal, as is the case, for example, in the multicellular organism Ascaris megalocephala.
Which of the three cited possibilities, allowed by our theory, is in fact realised in Pleodorina can, of course, be decided only by means of special experiments. For this purpose one would have to make use of the same methods of the mechanics of development which are applied in works with multicellular organisms. The application of these methods to colonial forms would be all the more desirable and interesting from the theoretical point of view in that here we have transitional stages to the phenomena of determination in multicellular organisms.
In discussing all the cited cases of reproduction in cultures and colonies of unicellular organisms, we proceeded from the assumption that the cells forming the cultures or colonies and arising by way of division represent independent living systems, dependent upon one another only in so far as they mutually influence each other. This interaction between the cells may be entirely lawful and very complex, but so long as we regard it only as the result of such interaction between systems essentially independent of one another, we of course have no right to regard the culture or colony of unicellular organisms as a whole as a living system, that is, we have no right to extend our general laws of motion of living systems to the culture or colony as a whole. The statistical regularities which we observe in them are conditioned by the laws of motion of the separate living systems of which they are composed, and by those purely external causal relations which exist between these systems. Therefore in these cases the causal-analytic method may prove sufficient (at least in principle) for the elucidation of the phenomena observed in cultures and perhaps some colonies, on condition that the laws of motion of the separate cells composing the system be taken into account. But when we regard the multicellular organism as a living system and strive to explain the phenomena observed in it as manifestations of the general laws of motion valid for every living system, then evidently an analysis of the interaction between its separate parts (cells) — that is, the causal-analytic method — is already insufficient. Then we must investigate to what extent and in what manner the observed phenomena represent manifestations, particular cases, of these general laws of motion. Then the interaction between the separate parts (cells) must itself follow from these general laws, that is, must represent forms of their realisation (let us here point also to what was said in Chapter II of Part I concerning historical regularities). We must be in a position to show that here too the same laws of metabolism, of the boundary of growth, of the fundamental process and of their relations manifest themselves, and that the interaction between the separate parts itself follows these laws. The same holds also for the phenomena of reproduction in multicellular organisms.
Every multicellular organism arises, as is known, by way of numerous divisions from a single cell. In unicellular organisms we had the theoretically simplest case: the system — the cell — attains its boundary of growth, whereupon there sets in the fundamental process, leading to division. If we consider that our general laws are valid also for multicellular organisms, then we arrive at the following apparent contradiction: the multicellular organism possesses a boundary of growth exceeding the magnitude of the initial mass (that is, of the initial cell) many thousands, or even millions, of times; while on the other hand we assert that the cell divides after it has reached its boundary of assimilation. The division itself leads to the restoration of the initial potential, whereas on the basis of our general laws we must consider that the potential constantly diminishes in consequence of growth.
In cultures of unicellular organisms we have already shown that, in consequence of interaction — that is, in essence, of external actions — certain cells, with a known statistical regularity, do not attain their natural boundary of growth and therefore produce daughter cells with a lowered potential or smaller mass. But what in unicellular organisms sets in as a chance realisation of the possibilities laid down in the system, in multicellular organisms takes place as a lawful manifestation of the system as a whole. In such a case the indicated contradiction is resolved in the following manner: the generative cell of the multicellular organism likewise possesses its boundary of assimilation, lying not much higher than in unicellular organisms; consequently, in this cell too the fundamental process must set in not later than the attainment of the boundary of growth, but here the fundamental process leads not to the formation of two living systems, but to the increase of the very same system; as a result of the repeated inclusion of the fundamental process there takes place a raising of the boundary of assimilation of the system. The mechanism of these repeated fundamental processes remains the same as in unicellular organisms: it is the division of cells. This can be shown graphically on our -diagram (Fig. 18): the boundary of growth of the egg cell would lie, if the latter were a unicellular system, at ; before its attainment there sets in the fundamental process (division) , leading to the formation of a greater mass with a lowered potential; further growth takes place along the curve , characterised by another parameter , as in metamorphosis (see Fig. 11), until the fundamental process again sets in, and so on. This diagram directly resolves the above-indicated contradiction. Along the curves of metabolism, growth beyond is impossible (at each curve of metabolism possesses a point of tangency with the corresponding curve of the basal metabolism and intersects in the sequel only the curves with a smaller free energy; see above). In consequence of the repeated inclusion of the fundamental process (–, –, and so on), however, a considerably flatter curve is obtained, whose point of tangency with the corresponding hyperbola lies, consequently, considerably further off, that is, the system thus arising possesses a considerably higher boundary of growth. On the figure this curve of metabolism is denoted by a dotted line. Thus we see that it is precisely by means of the non-utilisation of the boundary of growth of the cell that the possibility is created of an almost unlimited raising of the boundary of assimilation of the entire system. We see also that, with respect to metabolism, our general laws retain their force also for the multicellular system: we observe a gradual fall of the potential, accompanied by an increase of mass, and in consequence of this a boundary of growth, exactly determined by the course of the new curve of metabolism that arises. This difference will lie there where the new curve of metabolism makes contact with the corresponding hyperbola. Consequently, here in principle all the equations derived from our basic laws retain their significance.
Thus, from what has been said it follows that between cell division in unicellular and in multicellular organisms there is an essential difference. Whereas in unicellular organisms the fundamental process, upon which the process of division is based, leads ordinarily to the formation of cells with the initial potential, in multicellular organisms this fundamental process leads to the formation of cells with a gradually lowering potential, that is, with a lower boundary of growth. Unfortunately, we do not yet have sufficiently sensitive methods to measure directly the potential of cells and tissues. Neither the calorimetric methods nor the electrometric ones, mentioned in the chapter on the theory of living matter, are, it would seem, suitable for this purpose; more may perhaps be expected from the optical methods adduced there also. But if at the present time we cannot yet, by means of direct measurements, verify our conclusion concerning the gradual fall of the potential of cells in multicellular organisms, there nevertheless exist facts confirming this conclusion and lending themselves to explanation with its help. With respect to this question, one should point to the investigations of E. M. Vermel and his collaborators, devoted to the study of the dimensions of cells.[Author’s note:] Z. f. Zellforsch. u. mikrosk. Anatomie, Bd. 20, 1933.
In these investigations of the magnitude and growth of cells, Vermel arrives at the conclusion that “cells, as independent systems, are characterised by a greater magnitude than that which is manifested in the organism, or, in other words, the organism exerts a coordinating action upon the magnitude of the nuclei (cells) in the sense of establishing a definite limit to the increase of their dimensions”. By a series of comparative investigations on explanted tissues, on the cells of malignant tumours, and upon the application of various poisons, Vermel showed that here it is a matter not of the influence of various actions on the increase of the capacity for growth of the cells of the tissues of the multicellular organism, but of the fact that these cells, in tissue connection, did not attain that boundary of growth which they could attain outside the organism or upon disturbance of the correlative relations. It is not difficult to establish that these results directly confirm our conclusion that the cells of the multicellular organism do not attain their boundary of assimilation and divide before this.
Another conclusion which we can draw from what has been said lies in the fact that multicellular organisms do not consist of cells essentially equal in value to one another and to the egg cell, and possessing a different destiny and different potentials only in consequence of mutual influence upon one another. On the contrary, from our considerations it follows that the construction of the multicellular organism out of cells — that is, out of systems which can also exist more or less independently and possess a considerably smaller magnitude than the majority of multicellular organisms — that this construction is founded upon the fact that, without the fundamental process — that is, along a single curve of metabolism alone — living matter possesses limited bounds of growth. The limit of growth can be essentially raised only by means of the repeated inclusion of fundamental processes. This inclusion takes place in the same form as in unicellular organisms upon their reproduction, which, as we have shown, is likewise based upon the fundamental process. The principle of construction out of cells, characteristic of all living systems — this essentially morphological, general-biological law — thus appears as a consequence of our general laws of motion of living matter.
Thus, the considerable raising of the boundary of growth, arising in the course of evolution, as compared with its level in unicellular organisms, could take place only by means of the inclusion of fundamental processes into the processes of growth and development. As is known, Severtsov[Author’s note:] Morphologische Gesetzmässigkeiten der Evolution, Berlin, Verlag J. Springer, 1931. explains the appearance of new forms in the course of evolution, among other things, by the fact that certain new morphogenetic processes are included in the process of development. Therefore the division of cells must be regarded as the first of these inclusions in Severtsov’s sense.
Thus, our general laws of metabolism retain their significance, as we have seen, also for multicellular organisms, in consequence of the arising of a new curve of metabolism; it nevertheless remains for us to investigate how matters stand with the fundamental process and with reproduction in the new organism arising in this way. First of all it is clear that in it the fundamental process will no longer be confined to separate cells, as was the case in cultures of unicellular organisms, but will embrace the entire system; that this does indeed take place is proved by the above-mentioned experiments of Goetsch and Hartmann on the individual immortality of multicellular organisms, and also by the above-adduced observations on metamorphosis, which show that the rejuvenation, or the raising of the potential of one part of the organism, or the rebuilding connected with this, takes place at the expense of the dissimilation of another part of the organism. The same is shown also by the mentioned facts of the prolongation of life by means of the prevention of flowering in plants. Thus, in principle reproduction in multicellular organisms takes place in the same way as in the case of multiple division (Fig. 19), with only the difference that in multicellular organisms the curve of metabolism is not simple, but complex, as is shown on the figure. But this is only the simplest case in the fundamental respect. Probably it is realised in certain worms and plants, in which the formation of mature sexual cells (or flowers) directly leads to the death of the entire organism.
A considerable complication arises by virtue of the following circumstance: the germ cell with the free energy possesses its boundary of growth at , while the organism arising by means of the inclusion of fundamental processes — at , lying considerably higher, the order of magnitude of being smaller than the order of , that is, . But this means that the fundamental process may lead to the formation of products with the free energy — that is, full-value germ cells — long before the attainment of . We then obtain a lowering of the free energy of the organism as a result of the inclusion of fundamental processes leading to the formation of germ cells, although the boundary of assimilation is still far from being attained. This lowering is attained partly by means of a diminution of mass, partly by means of a lowering of the potential of the organism. In our diagram such a case can be depicted as indicated in Fig. 19. That is to say, during the growth of the multicellular organism, together with the fundamental processes included in it — the divisions of cells — the fundamental process sets in at and leads to the formation of germ cells at the expense of the lowering of all the free energy of the organism from to . This process may set in, as we see, at the most diverse stages, and moreover repeatedly or periodically. But in contrast to cultures of unicellular organisms, its onset in multicellular organisms is in each separate case secured in a definite manner. Thus, in multicellular organisms we obtain the following picture: all cells before the onset of the fundamental process are equal in value, that is, possess a potential lowered correspondingly to the number of divisions accomplished, while full-value germ cells are formed at the expense of the potential or mass of the remaining cells. Consequently, whereas in a culture of unicellular organisms a certain number of cells perish, in consequence of mutual and external actions, in accordance with the laws of chance, in multicellular organisms the formation of full-value germ cells is secured precisely by the perishing, or the more rapid dying-off, of other cells. We have already said that this fundamental process may set in at the most diverse stages. If it takes place during the first divisions of cells, then we have before us the so-called germ track.[Author’s note:] By this is by no means to be understood a complete separation of the development of the germ cells from the development of the organism as a whole (see below). This is a case analogous to the first case in cultures of unicellular organisms. There the “germ track” (Fig. 14) arose in such a way that at the least one of the full-value cells always has the possibility of dividing in good time, that is, after the attainment of the boundary of growth. Here, however, the first two cells inevitably possess a smaller potential than the first. Therefore the formation of a full-value cell is here possible only on condition that one of them gives up a part of its energy to the other, in consequence of which its boundary of growth is lowered. Then we obtain the same picture as in Fig. 15. But there it is the result of chance mutual and external actions, while here it is a lawful form of the development and of the fundamental process of the organism as a whole. If the fundamental process takes place later, then we obtain the same picture as in the second case in cultures of unicellular organisms: after a definite number of divisions there arise full-value germ cells and cells with a lowered potential. But whereas there the separate cells formed full-value ones by means of unequal division, while the rest perished in consequence of external conditions, here all the cells form a system and the formation of full-value germ cells is the result of the fall of the potential in the rest.
The independence of the systems with respect to the fundamental process in unicellular organisms, on the one hand, and their union in multicellular organisms, on the other, can be shown directly by way of experiment. Indeed, whereas in multicellular plants and animals the damaging of definite cells lawfully leads to a rebuilding of the nucleus and to division of the neighbouring cells, as is known from the works of Haberlandt on the so-called wound hormones, on the changes of the nucleus after wound stimulation, and also in general from the phenomena of regeneration, in unicellular organisms this action of the injury of one cell upon others cannot be attained: in them it takes place only within a single system, a single cell. The energy of equalisation of one part of living matter — that is, the structural energy — leads to its raising in another part only within the bounds of a single system. We see in this way that, with respect to both metabolism and the fundamental process, multicellular organisms are subject to our basic laws. But just as the basic laws of metabolism and growth are realised by means of a large number of included fundamental processes upon the divisions of cells, so too in the most diverse manner is the fundamental process realised also in the reproduction of the multicellular organism. Thus, the lowering of the free energy may take place by means of the partial or complete cytolysis of certain cells or groups of cells, by means of the lowering of the potential of some, or by means of an even lowering of it in all the cells. Further, in the fundamental process not all, but only some cells or groups of cells, acquire anew the initial potential. The latter, becoming subsequently germ cells, may be equal in value, with respect to potential, to the remaining cells and acquire their heightened potential at the expense of other parts of the organism in consequence of their position, structure, and so on; or else they may, already from the start, in consequence of the smaller number of divisions accomplished or the smaller fall of the potential during divisions, possess a higher potential than the remaining cells, as a result of which they more easily attain the initial potential in the fundamental process.
Indeed, we know that the future germ cells very often lag behind the remaining cells in their division. On the other hand, we know that in a number of animals (sponges, coelenterates) and plants the future germ cells lie isolated and scattered in the tissues. It must be supposed that such a distribution of them is the morphological expression of the fact that these cells, in dividing, attain their initial potential, that is, that they, like the cells of unicellular organisms, do not divide “prematurely”; these systems are thus more or less independent. But it would be an error to consider that such cells can be transformed into full-value germ cells without the fundamental process. Here it is only a matter of the fact that, in the case of a smaller number of divisions or of early-setting-in isolated “full-value” divisions (that is, after the attainment of the boundary of growth of the given cell), a smaller energy is needed for the restoration of the initial potential in the fundamental process. Even in the limiting case, when already the first division of the germ cell is unequal and consequently only one of the products of division is full-value and in the sequel divides as a full-value cell — even here we are compelled to accept that this unequal division is the result of the fundamental process of the entire system. A whole series of facts speaks in favour of the fact that in these cases too the fundamental process — that is, the lowering of the potential or the diminution of the mass of the organism and the raising of the potential of the germ cells at the expense of this structural energy — is necessary and does indeed take place. Thus, we know that the formation of mature germ cells — for example, upon the spawning of roe in fishes — is accompanied by a considerable cytolysis of cells in the tissues and by a lowering of the rate of growth. This phenomenon is equal in value to the inclusion of the fundamental process in metamorphosis.
Of still greater interest in this respect is the behaviour of the above-mentioned scattered meristematic cells, capable of reproducing and forming organs and whole organisms. They receive this capacity, however, only on condition of the damaging of neighbouring (or other) cells of the body, that is, through the fundamental process. Thus there finally becomes comprehensible also the fact that a separate cell of a tissue always loses its capacity to grow, divide, and form a full-value organism. In order that such a cell might preserve this capacity, it must either be a full-value, mature germ cell, having already received its initial potential at the expense of the energy of the remaining cells, or else together with it there must be present other cells, whose death subsequently renders possible the fundamental process and consequently the raising of its potential.
Chapter 3: Adaptation, Irritability
The general phenomena of life which we have hitherto investigated — metabolism, growth, and reproduction with their regularities — could be presented as particular cases of the development of the general laws of living matter. But in doing so we made throughout the assumption that these laws of motion can manifest themselves without hindrance, that is, that no such changes arise in the surrounding environment as would disturb the laws of motion of living matter. That a disturbance of these laws can set in only as a result of changes in the state of the surrounding environment, that is, only as a result of external influences — this follows directly from our assertion that these laws of motion are proper precisely to living matter itself, are inherent in it. Therefore the cause provoking their disturbance can lie only outside the matter forming the living system.
According to our principle of stable non-equilibrium, upon every change in the state of the surrounding environment the living system performs work directed against the onset of equilibrium under these altered conditions, that is, work serving to maintain the capacity for work of the system. But since the capacity for work, or the non-equilibrium, of the system is conditioned by the state of living matter, by its molecular structure, this means that upon every change in the state of the surrounding environment, at the expense of structural energy, work is performed directed towards the preservation or formation of living matter. We see that this work is accomplished in the form of the fundamental process and of metabolism; it is therefore not work in the physical sense, in which a system performs work against the surrounding environment, correspondingly lowering its free energy. But since the work necessary for the preservation of living matter depends upon the state of the surrounding environment, that is, is different under different conditions, whereas the capacity for work of a given system, that is, its structural energy, is limited, the work against the onset of equilibrium can be sufficient for the preservation of living matter only within known limits of change of the external environment; namely: if, under altered external conditions, more energy must be expended per unit time than can be formed as structural energy in the form of living matter, then under these conditions the mass, or the potential, of living matter will diminish, and the system will die off, since the work itself is indeed performed at the expense of the structural energy of living matter. Consequently, in this case a part of the free energy of living matter will no longer perform work against equilibrium, and the laws of motion of living matter will be disturbed. The property following from our principle — that within known limits the laws of motion of living matter are not subjected to disturbances under various states of the external environment — we call the adaptation of living systems. The conclusion, likewise following directly, as we have seen, from our law — that the work against equilibrium suffices for the preservation of the living system only within known limits of change of the surrounding environment — means that every living system possesses its own limits of adaptability.
It now falls to us to investigate by what these limits of adaptability are determined and what happens when we pass beyond them. How can these limits be determined, and upon what properties and states of living systems do they depend?
In order to give an answer to these questions, we must first elucidate certain points, above all such as concern the very concept of adaptation. The phenomenon of adaptation is contained, as we have already said, in our laws of motion of living matter, and therefore represents a property of all living systems. Adaptation is the dependence, expressed in the laws of motion, of changes in the state of living matter upon changes in the surrounding environment. In respect of direction and degree this dependence is determined by the laws of motion of living matter: the direction is determined by the requirement that the change of state should lead once again to a non-equilibrium state, or to the preservation thereof; and the degree by the fact that the work required for this can be performed only directly at the expense of the free energy of living matter, that is, at the expense of its structural energy. Thus we have a dependence of changes in the state of living matter upon the external environment, the regularity of this dependence consisting in the fact that the laws of motion are realised under various states of the surrounding environment in dependence upon the latter, in consequence of which there is preserved the state characteristic of living matter. The law-governed element in adaptation thus consists not in the fact that to each change in the state of the surrounding environment there corresponds a definite change in the state of living systems, but in the fact that within known limits all these changes in the state of the system lead to the preservation of one and the same characteristic state. What is essential for adaptation is not the various forms of manifestation of the laws of motion of living matter under various conditions, but the preservation of these laws themselves. Therefore, if under a lowered O₂ pressure cells respire less, or under a raised temperature more, intensively, this dependence does not yet signify adaptation. We designate this dependence as adaptation only then and only in so far as the system performs work which, in the first case under lowered, and in the second under raised, oxygen consumption leads to the preservation of the non-equilibrium state, that is, in so far as thereby the laws of motion of living matter, and consequently this matter itself, are preserved.
These, seemingly so simple and trivial, considerations are very often left out of account, in consequence of which the problem of adaptation and the regularities connected with these phenomena are excluded from biology. Indeed, if one does not bear in mind that the realisation of the laws of motion of living matter depends upon changes in the surrounding environment, and that the degree of this dependence is limited by these very laws, then we inevitably arrive at the conclusion that the laws of motion must be realised under any surrounding environment whatever, and always without change. Thus we arrive at the conceptions developed, for example, by Uexküll, according to which organisms and the inorganic surrounding environment are adapted to one another, that is, at something like a “pre-established Leibnizian harmony”. Living matter has its own laws, non-living matter too, but both are in correspondence and do not hinder one another.
But if one forgets that the essence of adaptation consists not in the dependence of changes in the state of living matter upon changes in the surrounding environment, but, on the contrary, in the preservation of its laws of motion, that is, of living matter itself with its characteristic structure and state, notwithstanding this dependence — if one forgets this, then the problem of adaptation likewise falls away, since such a dependence exists in non-living systems as well. But together with this, living matter, with its own laws of motion, then disappears of necessity, and we arrive at the conceptions of mobile equilibrium, of Le Chatelier’s principle, and so on.
In both these cases, as we have seen, the problem of adaptation falls away. In the first case it is replaced by an adaptedness given from the outset; in the second it is simply denied, together with the remaining special laws of living matter. But in both cases the existence of limits of adaptation remains unexplained and incomprehensible. If, however, we bear in mind our laws of motion of living matter, then we shall be secured against such errors. It is then clear to us that adaptation signifies not some special function, but the preservation, within known limits and in unaltered form, of the laws of motion of living matter, that is, of this matter itself; and the existence of limits of adaptability follows directly and of necessity from the fact that, on the one hand, for the maintenance of living matter it is necessary to expend work depending, in qualitative and quantitative respects, upon the surrounding environment, while on the other hand the capacity for work is, in quantitative respect, limited, since this work can be performed only at the expense of the structural energy of living matter.
Thus it falls to us to investigate the conditions under which the laws of motion can be realised without hindrance, and, proceeding from these laws, to elucidate those general conditions under which they will be disturbed.
The first difficulty upon which we here stumble lies in the question: when is a disturbance of the laws of motion of living matter present? The simplest thing would of course be to say that this takes place when living matter can no longer be preserved, that is, when the organism dies. But from what was said in the preceding chapters we know that the dying of individual organisms sets in inevitably even under the unhindered realisation of the laws of motion of living matter (though under the correct course of the processes living matter, as we have seen, is preserved by means of the fundamental process and the reproduction connected with it). Here, however, we shall not consider the question of adaptation and its limits in living matter in general, that is, including reproduction and the new generations arising as a result of it. This question will be considered further on, in the chapter on evolution. Here we shall confine ourselves to the problem of adaptation and its limits in individual organisms. From this it is evident that the answer given above to the question of when a disturbance of the laws of motion of living matter is present is incorrect.
We might attempt to give this answer a more precise form and to say that a disturbance of the laws of motion of living matter is present when the organism dies “prematurely”, that is, that the organism is not in a position to adapt itself to definite external conditions if, under these conditions, it dies earlier than would correspond to its “mean, normal” length of life. It is apparently precisely this conception that usually prevails among biologists when they speak of adaptation and its limits. In limiting cases it indeed gives rise to no particular difficulties. If we transfer an organism living for several years under its natural conditions into an environment deprived of oxygen or with a temperature of 60°, and the organism thereby dies within the course of several minutes or hours, then it is of course not subject to doubt that it could no longer adapt itself to this environment or temperature. But the unsatisfactoriness of this conception becomes immediately clear if we take into account that for each species of organisms there exist very various “mean, normal” durations of life corresponding to the conditions of the external environment in which they live or are cultivated. Thus, for example, from the works of MacArthur and Baillie cited in the chapter on metabolism it is evident that the normal length of life of Daphnia magna equals, at 8°, 108.4, and at 28°, 29.2 days; while, according to the data of Shcherbakov, Drosophila lives, at 18°, on average 78.7 days, and at 24°, on average 50.4 days. Which then of these magnitudes is to be acknowledged as the “normal, mean” length of life and taken as the criterion of the limit of adaptation? If we were to consider the high temperature already harmful, in so far as under it the mean length of life is shortened, then we should have to consider harmful those temperatures too at which these organisms live in nature, since there are lower ones at which they live considerably longer. We should then arrive at the conclusion that organisms are not adapted to their natural environment. Consequently we cannot simply speak of a normal, mean length of life. We have only the right to speak of a mean length of life under known conditions of the surrounding environment. But then the length of life cannot serve as a criterion of the limit of adaptation, since the very concept of adaptation and its limits is bound up with changes in the surrounding environment.
But the concept of the mean length of life under definite given conditions of the environment is likewise not entirely clear, even if these conditions be kept as constant as possible. It is a statistical magnitude, or one of the many statistical magnitudes worked out for the characterisation of the length of life in definite species, but the biological sense of which is unclear and perhaps even altogether absent. To introduce a statistical magnitude such as the mean length of life has, of course, sense only in the case where this magnitude, first, characterises a known property of the given collective, that is, in our case of the given species of organisms, and, secondly, if this property actually possesses a biological sense, a biological significance. But we have seen that the mean length of life, apart from its dependence upon external conditions, is in general not a characteristic magnitude. If, however, we take strictly constant conditions of the environment, or rather, conditions whose fluctuations do not pass known limits, then the individual variability of the length of life will, in any case, prove to be conditioned not by phenomena of adaptation, but by differences in hereditary properties. But then the mean length of life can tell us nothing about this variability.
With the fact that we do not know exactly what biological magnitude the mean length of life is properly to measure, or what biological significance it possesses, is bound up also the circumstance that there exist the most various concepts of the length of life. Thus one speaks of the mean, of the probable, of the normal or typical, and of the maximal length of life. All these magnitudes differ strongly from one another. For example, for man the mean length of life (that is, the summed length of life of all individuals divided by the number of individuals) equals 50–55 years, while the probable length (that is, that upon the attainment of which half of all individuals die, in other words, that at which the curve of survivors falls to half of its initial magnitude) lies between 60 and 65 years. The normal length of life, that is, that at which the annual coefficient of mortality attains its maximal magnitude, equals in man 70–75 years (if infant mortality, that is, the initial magnitude, be excluded); while the maximal length is an entirely indeterminate magnitude, in man in any case exceeding 100 years. Although, so far as we know, there are no special investigations on this question, it can nevertheless be asserted with confidence, on the basis of the available material, that all these magnitudes do not change proportionally under various external conditions, so that their relations may undergo essential changes even in the case where every variability of hereditary properties in the individuals of the given collective is absent, that is, if we are dealing with genetically homogeneous material.
Thus we see that the length of life alone can in no way serve as a measure in the determination of the limits of adaptability, that is, on the basis of the length of life of a given organism we cannot decide whether a disturbance of the laws of motion of living matter was present under the given external conditions or not. If the criterion for the determination of whether some influence or state of the surrounding environment exceeds the limit of adaptation of the given organism is whether in such a case a disturbance of the laws of motion of living matter takes place, then we must of course find this criterion by proceeding from the laws of motion themselves, and not from some arbitrarily taken phenomenon of life. Since we here consider the question of adaptation and its limits not in living matter in general, but in individual, given organisms (see the introduction to the second Part), that is, the question of so-called physiological adaptation [физиологическое приспособление — fiziologicheskoe prisposoblenie], we must seek the criterion of the limits of adaptation in the disturbance of those laws of motion which follow from our general laws for the individual cycle of living systems; the laws relating to reproduction we must not here take into account. If we were to include also the regularities of reproduction with their disturbances, then we could not in general speak of a physiological limit of adaptation. The significance of this delimitation, however, is more profound; it will reveal itself with particular clarity further on, in the chapter on evolution, though even here it is already justified by all that has been said above. The laws by which the individual cycle proceeds are given, as we saw earlier, by the very laws of metabolism, as they follow from our general laws of motion. The processes of reproduction, on the contrary, are founded upon the laws of the fundamental process. Consequently, we have full right to investigate the limits and results of the disturbance of the laws of metabolism separately from the laws of the fundamental process, although in the chapter on reproduction we did establish a law-governed connection between the disturbance of the processes of metabolism and the fundamental process: the latter always sets in in the case where metabolism is disturbed or can no longer be realised in consequence of the potential of the living mass diminishing in the course of ageing.
Thus, in this regularity we have, apparently, the sought-for criterion of the limit of adaptation. This criterion is formulated as follows: a disturbance of the laws of motion in consequence of changes in the state of the external environment is present in the case where the latter provoke the fundamental process. We shall now occupy ourselves with the investigation of how far this criterion is suitable and what its sense is, that is, what it properly signifies.
First of all, let us adduce several examples. Let us take a unicellular organism in which unfavourable external conditions (temperature, nutrition) have provoked the formation of cysts, founded upon the fundamental process. In this situation the following is contained: at the expense of a part of its structural energy, which under other conditions the cell could have employed for the continuation of metabolism, it has formed a cyst with a higher potential. The laws of metabolism were consequently disturbed, the latter was prematurely terminated as a result of the external conditions. According to our criterion, we consequently can no longer regard the formation of the cyst as a physiological adaptation. Another example: in a newt a limb is cut off, which subsequently regenerates. According to what was set forth in the preceding chapter, regeneration too is partly connected with the fundamental process. A group of cells was damaged, the latter could not employ their structural energy for the continuation of metabolism, metabolism was prematurely interrupted, and this energy went in part to the raising of the potential and to the formation of structures in the regenerating cells of the meristem. On the basis of our criterion, regeneration consequently likewise cannot be considered a physiological adaptation, that is, the external influence in the form of an incision with a knife exceeds the limit of physiological adaptation. A third example: a unicellular organism is transferred into an environment with a higher temperature and begins to divide more rapidly. In this case the external conditions provoke premature division, that is, properly the premature onset of the fundamental process, since division is indeed founded upon it. Therefore, at first sight we should have to say that every temperature provoking accelerated division of cells already lies beyond the limits of physiological adaptation. But then all temperatures below which there are still such as prolong the period of division without killing the animal would lie beyond the limits of physiological adaptation, and we should arrive at the same conclusion as with the criterion based upon the length of life, since the latter would coincide with our new criterion. Consequently, in this case we can no longer, without reservations, put into this criterion the same sense as before, and say: the cell has formed two new cells with a higher potential at the expense of a part of its structural energy, which under other conditions it could have employed for the continuation of its metabolism; the laws of metabolism were disturbed, the latter was prematurely interrupted as a result of the external conditions. We cannot assert this, since it is known to us that metabolism is accelerated at a high temperature, so that the potential has perhaps already fallen earlier to such a magnitude at which metabolism could not continue, and the fundamental process, connected with division, has set in — earlier indeed in respect of time, but not prematurely in respect of the laws of metabolism. In other words, metabolism was not interrupted, but attained its normal end. In just the same way one can artificially attain an early end of metabolism by means of other influences provoking a more intensive metabolism, by way of accelerating the processes of dissimilation, or, for example, by way of compelling the formation of certain new structures. The latter sets in, for example, in the case where, instead of provoking regeneration as in the second example, we influence (as is often done in developmental mechanics) the normal formation of certain structures, for example by way of pressure, in consequence of which structures are formed which would otherwise not have arisen. Consequently, the last two cases are characterised by the fact that the external influence itself provoked not the fundamental process, but only a raising of the intensity of metabolism, in consequence of which an earlier end of the latter set in. But this end sets in, in such a case, at the same state of living matter, that is, at the same magnitude of the potential, as in the norm. Consequently, here no disturbance of the laws of metabolism took place. In such cases we say that the external influence lay within the limits of physiological adaptation. We think that these examples sufficiently elucidate the biological sense of the criterion of the limits of physiological adaptation derived from our general laws. This criterion runs: fundamental processes provoked by external influences, and the formations of new structures conditioned by the latter (cysts, regeneration, and so on), do not belong to physiological adaptation and represent manifestations of living matter which are characteristic precisely of the passage beyond the limits of physiological adaptability. From this same criterion it follows, conversely, that so long as the laws of metabolism are not disturbed, that is, so long as the whole structural energy is expended upon the
conduct of metabolism, upon the maintenance of the non-equilibrium of living matter at the expense of assimilatory work, or leads to the formation of germ cells along a hereditarily established path — so long are we still within the physiological limit of adaptability, independently of the course of the processes in time and of the special structures formed thereby.
Since we proceeded from our laws of motion, and specially from the laws derived for metabolism, it becomes at once clear to us that the course of the processes of metabolism in time, and consequently the length of life, cannot give us the criterion we require, since these laws say nothing about the course in time, that is, about the intensity of metabolism. The latter depends in great degree upon external conditions. But of course the velocity or intensity of metabolism influences in a decisive manner the moment of attainment of that potential at which metabolism cannot continue, or at which, in the hereditarily determined order, the fundamental process sets in, leading to reproduction, to the formation of germ cells. Nor is it of significance what special structures are thereby formed, so long as their formation proceeds at the expense not of the fundamental process but of assimilation, and so long as these structures are “full-valued”, that is, are preserved at the expense of their own structural energy by way of assimilation.
The criterion thus obtained, derived directly from the connection described in the preceding chapter between the metabolisms and the fundamental process, is unquestionably a suitable and sufficient criterion. It clearly shows us the biological sense of what we are to understand by physiological adaptation and its limits. At the same time there is contained in it the assertion that a change in the state of the surrounding environment exceeds the limits of physiological adaptation only then, if it leads directly to the destruction of living matter, since the fundamental process can set in only at the expense of a diminution of the living mass and its structural energy. But this is also the cause of the fact that this criterion, although it is correct and sufficient, is not necessary. For if the fundamental process can indeed be provoked only by way of the destruction of living matter, yet not every destruction of living matter inevitably leads to the fundamental process. We can, for example, subject the cells which in the first of the adduced examples formed cysts under the influence of temperature, nutrition, to the action of an altered temperature so rapidly that they are destroyed and do not form a cyst. Or we can destroy the limb or tissue of an organism incapable of regenerating, and then neither the fundamental process nor regeneration sets in. Here it is of course entirely clear that metabolism is thereby disturbed, that is, as a result of the external influence is prematurely interrupted, before the potential is so far lowered as to condition the death of the organism or the onset of the fundamental process and division at the hereditarily determined moment.
If thus the above-adduced criterion is indeed sufficient, then the necessary criterion of the limit of physiological adaptation is the disturbance of the laws of metabolism, derived from the general laws of motion of living matter. Consequently, we must find the criterion of the disturbance of these laws. Such a criterion follows directly from our theory of living matter and the laws derived from it in the chapter on metabolism. From these laws it follows that by the initial potential µ₀ and the initial mass m₀, and also by the magnitude, depending upon the given particular structure,
there is unambiguously determined the number of molecules which can be assimilated both during growth and after the attainment of the boundary of assimilation. Since the magnitude c itself depends upon the kind of assimilated molecules, the quantity of the latter will depend upon the kind of nutrition, but with a definite food it will be determined unambiguously.
All this is correct on condition that 1) the processes of metabolism proceed without disturbances and 2) the potential falls to a magnitude determined for each organism, after which the fall ceases. If we take this magnitude as equal to, or close to, zero, then the second condition falls away. The quantity of heat formed in the course of the whole of life per unit mass upon the fall of the potential from its level µₙ to zero, as a result of the work necessary for the metabolism of substances, that is, for the assimilation of the food molecules — this quantity of heat we call Rubner’s constant [рубнеровская константа — rubnerovskaia konstanta].
Thus, the unambiguous criterion of the physiological limit of adaptation is Rubner’s constant. So long as the magnitude of the heat actually formed equals Rubner’s constant, we are within the limits of physiological adaptation. If it is less, then we have before us a disturbance of the laws of motion as a result of the destruction of living matter by external influences, which leads either to the fundamental process or to the dying-off of a part or the whole of living matter. This criterion is unambiguous and, just like the first, includes not only the cases of change of the processes of metabolism in time, but also the cases of formation of new structures during development as a result of external influences, if this formation is not provoked by the dying of living matter. For even if such structures do require for their formation more work than those arising under other conditions, yet in what follows this surplus of structural energy will, under undisturbed metabolism, be given up again after the attainment of the boundary of assimilation, when the organism dies or these structures age as a result of their metabolism.
In general, for any value of the external conditions (temperature, pressure, oxygen content, concentration of known ions, and so on) and for each organism with identical initial magnitudes (µ₀, m₀, c), that is, with identical hereditary properties, we shall obtain, within known limits characteristic of the given conditions and of the given organism, an identical magnitude of Rubner’s constant, whereas beyond these limits the magnitude will be less. For some conditions the range of these fluctuations may be very inconsiderable, for others very great. As a limiting case of this regularity one may regard anabiosis, which is attained in some organisms by means of low temperatures or gradual desiccation. Both these factors act in the sense of a lowering of the intensity of metabolism; if thereby no dying-off of living matter, and consequently no disturbance of metabolism, sets in, then the length of life too must correspondingly increase; and if the intensity of metabolism thereby tends to zero and the length of life to infinity, then the product of both magnitudes too will approach a definite, finite magnitude. In other organisms the same conditions, upon attaining a definite limit, already provoke damage to living matter and thereby a disturbance of the processes of metabolism, which leads to the fundamental process and the formation of cysts, that is, the value of Rubner’s constant is here not preserved without limit. Finally, changes in the same external conditions (lowering of temperature and of humidity), especially if they take place very rapidly, may lead to damage to living matter without provoking the fundamental process, and we again obtain a magnitude smaller than Rubner’s constant. Experience shows us that the limit of physiological adaptation in relation to some factor depends not only upon the magnitude of the changes of this factor, but also upon the velocity with which they take place. Thus, in this case the damage to living matter is conditioned not by the new external conditions themselves, but by the rapid change of the latter. Therefore we have no right to say that a given temperature, a given pressure, degree of humidity, or oxygen content lies beyond the limits of physiological adaptation. But the matter naturally stands thus, that if the damage is provoked by rapid changes of some condition, it too takes place very rapidly, that is, it has already set in upon the new surrounding environment, and is not provoked by it. For example, if we suddenly transfer a freshwater organism into a high concentration of salts, then the damage will set in not in consequence of the sojourn in this concentration, since on occasion the very same organism, if it be transferred gradually, would remain unharmed in the very same concentration, that is, would form new structures or pass over to a more intensive metabolism, or both together, but without disturbance of the processes of metabolism, that is, without a diminution of the magnitude of Rubner’s constant. On the other hand, for the same organism there exists an upper limit of the concentration of salts, at which, independently of the velocity with which it is attained, damage takes place and, consequently, a diminution of the constant. Therefore we must make a distinction, at the least, between adaptation and its limits in relation to certain constant external conditions and in relation to changes of these conditions and their velocity. Whether the damage was provoked by the velocity of the changes of the given condition or by a more or less prolonged sojourn in the new surrounding environment, this can be decided in each particular case by means of an immediate return of the conditions to the initial state: if the damage appears in this case too, then it was provoked by the velocity of the change; if, however, there is no damage in this case, but it sets in upon a more prolonged sojourn in the unaltered new environment, then it was this environment itself, and not the velocity of the change, that provoked it. Experience shows that damage in consequence of the velocity of change, and damage in consequence of sojourn in definite, unchanging external conditions, are to a great degree independent of one another, that is, a prolonged action of a definite pressure, temperature, electromotive force, and so on, may provoke damage, whereas a very rapid change of these influences up to the same magnitudes by itself, that is, if they are immediately removed, exerts no harmful influence, and conversely.
This far-reaching independence, in the sense of harmful biological action, between the velocity of change of external conditions, on the one hand, and the prolonged action of the same conditions above or below known limits, on the other, is conditioned by the fact that, in respect both of the influence and of the reaction of living matter, we are dealing with two different things. As regards the differences of influence, here we must be in a position, in so far as we are dealing with influences of non-living nature, to establish this difference by physical methods, that is, we must have the possibility of distinguishing both kinds of influence by means of physical concepts. As regards the difference in the reaction of living matter to these two different influences, it is determined by the laws of motion of this matter.
Let us first elucidate the difference in influence by the example of pressure. When we investigate adaptation to prolonged influences by various pressures, we leave each of them constant for a more or less prolonged time. If some force, for example pressure, acts upon a body capable of displacement with mass m, then, as is known, this body receives an acceleration proportional to the force. But if the pressure acts in the form of an impact, that is, very rapidly changes from zero to a definite magnitude and then again quickly falls to zero, then, as is known, we are dealing with an impulse [импульс — impul′s], defined by the integration of the force over time, and the body receives a velocity proportional to the impulse. This shows that in the case of the action of a constant pressure and of a very rapidly changing one (an impact), two different magnitudes are decisive: in the first case the force, that is, the pressure itself, and in the second the impulse; the measure of action too is different: in the first case it is the acceleration imparted to the body, in the second the velocity. From this it follows that in both cases two different physical magnitudes are decisive. But it is more important for us to consider the case where the force, that is, the pressure or the pull, provokes a change in the state of the system upon which it acts. If, for example, a known pressure or pull acts upon an elastic spring, then the latter undergoes a change in its length, proportional to the magnitude of the force. The spring receives a potential energy corresponding to the force, in consequence of the deformation. If we provoke a rapid and transient change of pressure or pull, that is, if the spring receives an impulse, then the latter will disturb the equilibrium state, and the spring will begin to oscillate about this state, in order thereafter to establish it once again. Consequently, the constant action of a force here provokes a displacement of the equilibrium position, while a rapid and transient one — that is, an impulse — provokes not a displacement of the equilibrium position, but oscillations about it. In the first case the displacement of the equilibrium position or the potential energy of the spring may serve as a measure of the force; in the second the amplitude of the oscillations may serve as a measure of the impulse. But both the one and the other are valid only within known limits. If the pull exceeds this limit, then it will lead to a non-elastic deformation, and then the displacement of the equilibrium position or the potential energy of the spring can no longer serve as a measure of the force. The impulse too may, if it exceeds a known magnitude, lead to a rupture of the spring. Thereby the magnitude of that pressure at which the elastic deformation proves to be exceeded stands in no connection whatever with the magnitude of the change of pressure in the impulse, since in the latter what is decisive is precisely the integral over time.
What was said of the spring relates also to a gas situated under various pressures. To each pressure there will correspond, as an equilibrium state, a definite volume. The impulse, on the other hand, will provoke a vortical motion of the gas, after which very quickly the former equilibrium is re-established.
To the concept of the impulse in mechanics there is analogous the concept of the impulse of current () or the impulse of voltage () in the doctrine of electricity. When a current of definite intensity passes for a more prolonged time through a semiconductor, then there soon sets in a stationary state, an equilibrium of flow, whereby per unit time through the cross-section of the conductor there passes one and the same quantity of electricity, so that no accumulation of it takes place. But if we provoke a very rapid change of the strength of the current lasting only a short time, that is, if an impulse of current acts upon the conductor, then a new equilibrium will not set in, but a disturbance of the equilibrium distribution of charges in the conductor will take place, after which the former equilibrium too will soon be re-established. The same relates also to the prolonged action of a definite temperature or to a transient temperature change.
Thus we find, as a general rule, that a very rapid and transient change of some magnitude of the state of the surrounding environment, and a prolonged action by factors expressed by these magnitudes at the various values which they may have, differ from one another also in the physical sense, and must possess a different physical action. In the first case we are dealing with impulses, provoking up to known limits disturbances, but not displacing the equilibrium; in the second case, on the other hand, up to a known limit the equilibrium is displaced, but no disturbance is provoked. If this limit is exceeded, then in the first case (impulses) a displacement of the equilibrium is added to the disturbance, while in the second (prolonged actions) a disturbance of the equilibrium is added to the displacement of it.
Having made clear to ourselves the difference in the physical influence of the prolonged action by factors expressed by various values of the magnitudes of the state of the environment, on the one hand, and of the velocity of change of these magnitudes, on the other, we must, on the basis of the laws of motion of living matter, elucidate the changes of state and the conditions for damage to this matter under various forms of influence.
Upon the prolonged action of some factor of the environment on a living system we are, as has been said, dealing not with the onset of a new equilibrium corresponding to the altered environment, but with the preservation of the work-capable non-equilibrium state characteristic of living matter at the expense of the work performed by the free energy of the system itself. For this there is necessary, in dependence upon the conditions, a more or less intensive work. In such a case we speak of adaptation to the new environment. If the work required for adaptation to the given conditions must be, per unit time, greater than what the structural forces of the system can yield, then the capacity for work of the system can no longer be fully utilised, and gradually a disturbance of the laws of metabolism will set in; and since the work serves precisely for the maintenance of these structural forces, living matter itself will also be damaged. In this case we speak of the limit of adaptability.
If we have before us a rapid change of a magnitude of state, that is, an impulse acting upon living matter, then it temporarily disturbs the non-equilibrium state subject to preservation by way of a direct disturbance of the processes of metabolism maintaining this state. The state of living matter approaches the equilibrium one, that is, its structural energy diminishes. But since the disturbance was provoked by an impulse, the process of metabolism is immediately restored and the former non-equilibrium state again sets in. In this case we speak of stimulation [раздражение — razdrazhenie] by the surrounding environment. If the impulse exceeds a known magnitude, then equilibrium too may set in, and then metabolism can no longer be restored. To the disturbance of the non-equilibrium state of living matter there is added also a final disturbance of metabolism.
Thus, in adaptation we have before us, in the first place, a change of the intensity of metabolism and sometimes the formation of new structures; and if known limits of the altered magnitudes are exceeded, then as a result of the prolonged action there is added to this, secondarily, a disturbance of the state of living matter as well.
In stimulation we are dealing, in the first place, with a disturbance of the state of living matter, to which, if known limits are exceeded in consequence of too strong a disturbance of the state, there is added secondarily a final disturbance of metabolism as well, and consequently a dying of the tissue.
Before occupying ourselves with the mechanism of the disturbance of the state of living matter provoked by a stimulating impulse, we shall investigate, on the striated muscle as a typical example of an irritable tissue, these differences in the changes provoked by the action, on the one hand, of constant magnitudes, and on the other, of impulses.
If we act upon a striated muscle, for example the gastrocnemius or sartorius, by means of a constant stretching, then, on the basis of what has been said, we must expect a change of the intensity of metabolism, the state of the tissue remaining work-capable. If the stretching is too strong, or the action lasts too long, then after a certain time there must be added to this altered intensity of metabolism a disturbance of the non-equilibrium state itself. That this indeed takes place was shown by Feng[Author’s note:] Feng, J. of Physiol., vol. 74, no. 4, 1932. by means of direct measurements of oxygen consumption and heat formation.
The action of stimulation in the form of a contraction of the muscle, that is, a change of state, we obtain, on the contrary, by means of an impulse, that is, a rapid change of the force of stretching. That the disturbance of the state is attained thereby precisely by the impulse, and not by the magnitude of the acting force, is seen best of all from the fact that the reaction to the stimulation, that is, the contraction, sets in always after an, albeit very short but measurable, time after the impulse. For every stimulation a so-called latent period is characteristic. This precisely proves that the change of state is a consequence of the disturbance provoked by the impulse, and not of the action of the surrounding environment. The same, as is known, takes place also in electrical stimulation. Here too an impulse of current acts, and not a constant current. We obtain a contraction of the muscle only upon the making or breaking of the current, and not upon its prolonged passage. On the contrary, in the latter case the initial state is again re-established notwithstanding the further passage of the current, while the contraction sets in only after a known time after the impulse.
We shall now investigate the conclusions which follow from our principle of stable non-equilibrium, of the work of the structural forces, and from our theory of metabolism and of living matter, for the case of the disturbance by an impulse of the state of living matter and of the processes provoked by it, that is, the phenomena of stimulation.
In doing so we shall begin with the results obtained by experimental means on the striated muscle, and then investigate how far they have significance for smooth muscles and other irritable tissues.
In order to obtain a correct conception of the physiological behaviour of the striated muscle, we must proceed from an analysis of the state and metabolism at rest. In doing so we proceed from the general biological principle, according to which a living system is not thermodynamically in equilibrium, and the whole energy liberated in the spontaneously proceeding processes supplying energy is converted into work against the equilibrium which is to be expected under the given conditions. Applying this principle to the striated muscle, we must, consequently, assume that the processes supplying energy which proceed in a muscle under the action of its own or of a suspended weight (that is, the metabolism of this muscle in the state of rest) serve for the maintenance of the non-equilibrium state under the given load. A non-living, elastic body — a spring or a rubber band — will stretch under the influence of a load to such an extent until the force pulling downwards equalises with the force of tension, that is, until both forces acting in opposite directions balance one another. For the maintenance of this state there is no need for processes supplying energy. In a living system, such as the striated muscle, this equilibrium does not set in, the tension and the weight do not balance one another, the former being always greater than that magnitude which would correspond to the given weight. Consequently, this system corresponds, for example, to a spring which we stretch (and hence tense) more than would correspond to the suspended weight, say, by fixing it in this position. The work which we must in such a case perform from without, in order to preserve the system in this state, is performed in the living muscle by the energy-supplying processes of equalisation, that is, by the resting metabolism. The muscle tensed by the weight is, consequently, not simply a system with greater potential energy, like a spring stretched by a greater or lesser weight in equilibrium, but it possesses a greater free energy, which would diminish under the given load if this state were not constantly maintained by the energy-supplying processes. From this it follows that if these energy-supplying processes are switched off or can no longer take place, then the muscle must become shorter, and external work can already be performed only at the expense of a diminution of the free energy of the system, that is, at the expense of the spontaneously proceeding process of equalisation. Thus there is explained without difficulty the fact that the muscle contracts when it dies, and that, for example, a muscle poisoned with monoiodoacetic acid always falls into a state of prolonged contraction when the immediate source of energy — phosphagen — is exhausted.
But this same fact explains yet another fact well known in the physiology of the striated muscle: this is the observation that the capacity for work of the muscle, measured by the work at maximal stimulation, rises up to a known limit with an increase of the load. From the fact that the muscle possesses a greater tension than would correspond to the load, and that the energy-supplying processes of metabolism at rest always perform work serving for the preservation of this surplus of tension (work against equilibrium under the given load) — from these two circumstances it follows that the intensity of resting metabolism must rise upon an increase of the load, and that the switching-off of the energy-supplying processes will condition a greater work, since the process of equalisation will begin from a higher level. Of course, this can take place only up to a known limit, namely only so long as the source of energy or the intensity of resting metabolism is still in a position to secure the surplus of tension. If the load is increased still further, then a lowering of the capacity for work must set in, the more so since then the mechanical action of the weight itself begins directly to disturb or damage the system.
Thus, the first conclusion which we draw from our above-adduced premises is the following: the intensity of resting metabolism, or of the energy-supplying processes in the state of rest, must, up to known limits, rise with an increase of the load, and upon a further increase of the latter must fall; the load at which the maximum of the capacity for work of the muscle is attained must correspond to that at which the intensity of resting metabolism attains its highest point.
To verify this first conclusion, we took, as a measure of the intensity of resting metabolism, the current of rest, which is easy to register and which is the expression of the intensity of the whole aggregate of processes of metabolism in a resting living muscle, but in a dead muscle disappears.[Author’s note:] These experiments were carried out together with my collaborator V. A. Muzheev.
The experiments adduced in the chapter on the theory of living matter showed that the current of rest is conditioned not by the properties of membranes or boundary surfaces, but by the deformed, non-equilibrium state of the molecules; according to these experiments the current of rest is a measure of the deviation from the equilibrium state, and consequently of the intensity of metabolism too, in so far as this deviation is conditioned by the latter. That we here are not dealing simply with the registration of a distribution of ions on boundary surfaces in consequence of the selective permeability of the latter, is seen from the fact that the current of rest exists also in smooth muscle, where the absence of selective permeability for cations and anions has been proved.
The object of our experiments was the gastrocnemius of the frog, the distal end of which we damaged by immersion in hot water, after which the muscle was suspended in a moist chamber. Then we, in the usual manner, by means of non-polarising electrodes and a mirror galvanometer, led off from it the current of rest under various loads. The scale upon which the reflected beam fell was situated at a distance of 1½ m from the mirror of the galvanometer. The current of rest was observed over the course of several hours. With each load a series of numerous experiments was carried out, the results of which agreed well with one another. In Fig. 20 the results of these experiments are given, the time being plotted on the abscissa and the magnitudes of the current of rest, in percentages of the initial magnitude, on the ordinate. The curves for each separate load, or the points relating to the latter, are mean magnitudes derived from a large number of experiments with one load. The weight was suspended when the current of rest remained without change over the course of 5 minutes. The arrow indicates the moment of loading.
From the figure it is evident that the experiments gave the following result: the current of rest of an unloaded gastrocnemius gradually falls with time and over the course of 5 hours drops approximately to half of its initial magnitude. Under a load of 20, 40, 80, and 160 g, on the other hand, the current of rest exhibits an immediate rise at the moment of loading, then a further rise over the course of 10–20 minutes, after which a slow fall sets in, however less steep than the rise; and, in any case, the current of rest in the loaded muscle remains over the course of several hours considerably higher than in the unloaded one. While the current of rest of the unloaded muscle after 130 minutes amounts to only 65% of the initial magnitude, we have, for a load of 20, 40, and 80 g, after the same time, on average still 85%. A steeper fall is exhibited in the second hour by the muscle with a load of 160 g. This too speaks in favour of the view that the current of rest registers the energy-supplying processes in the state of rest; and in so far as we are dealing with a surviving, isolated muscle, deprived of the supply of nutrition, the source of energy is naturally limited, so that the more intensive was, in the first hours, the work serving for the maintenance of the state, the sooner this source is exhausted, and in what follows a more rapid fall must set in.
That this course of the curve is indeed conditioned by the processes of metabolism of the undamaged part of the muscle, and not, as is often thought, by changes in the boundary zone, we were able to show by clamping the muscle above this boundary zone and loading only the upper half by means of a thread thrown over a fixed pulley. The course of the curve remained, under these conditions too, the same.
Thus, we see that the current of rest increases up to a load of 160 g. But if we take a load of 400 g, then we see that only at the moment of loading is an inconsiderable rise observed, after which a sharp fall of the current of rest begins. The latter already after 30 minutes lies below the normal curve, and after 100 minutes amounts to less than 30% of the initial magnitude.
Thus, the first part of our conclusion is fully confirmed. Up to 160 g the current of rest increases, while at 400 g it already falls. On the basis of the above-adduced considerations, the maximal work of the gastrocnemius at maximal stimulation without support must on average lie between 160 and 400 g. Usually, however, the maximal capacity for work of this muscle is found at a load of approximately 50 g. But this figure relates to contractions with support, whereas in our experiments the muscle was loaded without support. Therefore we determined the maximal lifting work of the gastrocnemius without support and found that it always lies at a load of approximately 200 g.[Author’s note:] The determination of the maximal lifting work is carried out, as is known, by measuring the height of lift at maximal stimulation and multiplying this magnitude by the lifted weight. We see thus that the second part of our conclusion too is confirmed: both the current of rest and the capacity for work have their maximum at approximately one and the same load (a more exact determination of this limit, of course, has no sense in view of the individual fluctuations of separate muscles). The next question consisted in whether the current of rest does indeed depend upon the intensity of resting metabolism, and which precisely of the energy-supplying processes is responsible for it in the first place. In order to exclude completely even very weak oxidative processes, we poisoned the muscle with KCN and found that the current of rest does not change noticeably as a result of such poisoning, since on average the strength of the current was identical in the poisoned and the unpoisoned muscle, and its changes in time under the influence of various loads exhibited the same picture.
An entirely different, characteristic course of the current of rest we find in a muscle poisoned with monoiodoacetic acid. First of all, it was striking that the initial magnitude of the strength of the current of rest in a muscle poisoned with this acid usually lay considerably higher than in an unpoisoned one. For the most part, in a muscle poisoned with monoiodoacetic acid the galvanometer exhibited an initial deflection exceeding almost twofold the deflection in an unpoisoned muscle. Considerably higher too in the poisoned muscle was the percentage rise upon loading. In Fig. 20 there is depicted the course in time of the percentage changes of the current of rest in a muscle poisoned with monoiodoacetic acid under a load of 40 g. We see that as a result of this load the current of rest rises by almost 50% of its initial magnitude, whereas in the unpoisoned muscle the rise under the same load amounted to less than 10%. On the contrary, we see that the fall of the current of rest in the last two hours takes place most rapidly of all in the poisoned muscle and, crossing all the curves up to and including 160 g, at the end of the third hour falls to 45% of the initial magnitude.
The stronger current of rest, and also its greater increase upon loading and the steep fall in what follows, in a muscle poisoned with monoiodoacetic acid show that under poisoning with this acid the energy-supplying processes, serving for the maintenance of the surplus of tension upon loading, here proceed considerably more intensively, and the limited source of energy is exhausted sooner. But since we know that monoiodoacetic acid provokes a retardation of the formation of lactic acid and a more rapid, unhindered decomposition of phosphagen, we must assume that the intensity of the current of rest depends upon the intensity of the decomposition of phosphagen and is proportional to it. Just as the working muscle poisoned with monoiodoacetic acid loses its capacity for work sooner in consequence of the more rapid decomposition of phosphagen and the absence of its restoration, so too the loaded poisoned muscle exhibits a stronger current of rest and a more rapid fall of it.
There exist, however, as we have already mentioned, also direct measurements of metabolism or of heat formation upon loading without contraction; such measurements were made by Feng, and his results fully correspond to the considerations and experiments adduced here. In his work on the thermoelastic properties of muscle[Author’s note:] Feng, The thermoelastic properties of muscles, J. of Physiol., vol. 74, no. 1, 1932. Feng found that upon loading the m. sartorius of the frog up to 20 g a positive heat formation is on average observed, while above 20 g a lowering of heat formation sets in. Feng in his work endeavours to explain this effect independently of the processes of metabolism, as a thermoelastic effect. He therefore assumes that this reversal of the thermal effect at a greater load is conditioned by a negative heat formation in the connective tissue upon tension, which at a greater load overrides the positive heat formation of the muscle fibres. That this explanation is untenable is shown already by the fact that the raised heat formation upon loading, just like the greater intensity of the current of rest, may last more than an hour, whereas the thermoelastic effect, for example in a dead muscle, is not only considerably less even upon prolonged loading, but, over and above this, disappears immediately, as was established by my collaborator V. A. Muzheev in the Leningrad laboratory of Professor Kupalov. On the contrary, the reversal of the thermal effect above a definite load, found by Feng, is explained without difficulty by our
above-adduced conceptions, according to which above a definite load the muscle can no longer create the necessary surplus of tension, while the load itself already disturbs the structure and retards the processes of metabolism, whereby is explained the diminution of the maximal capacity for work of the muscle and the lowering of the current of rest.
To the recognition of the incorrectness of his explanation Feng was bound to come himself on the basis of another of his works, published in the same number of the same journal. Here he directly investigated the formation of heat and the consumption of oxygen upon loading, and found a prolonged intensification of metabolism under the action of the weight. In this work Feng arrives at the same conclusion to which we too arrived on the basis of the above-adduced considerations and observations on the current of rest, namely, that perhaps a known level of phosphagen content is necessary in order that a noticeable intensification of resting metabolism upon loading should set in. This probable conclusion Feng apparently bases upon the observation according to which a muscle poisoned with monoiodoacetic acid exhibits a raised heat formation, but the latter, once lowered, can no longer be restored even in oxygen. But Feng writes that the reaction to loading remains undiminished upon poisoning with monoiodoacetic acid (“the stretch-response exists undiminished in muscles poisoned with iodoacetic acid”); we, on the contrary, find in the poisoned muscle, correspondingly to the unhindered, more rapid decomposition of phosphagen, a considerably greater intensification of the current of rest upon loading. But if we compare Figs. 7 and 8 in Feng’s work, then we see that the unpoisoned sartorius under a weight of 30 g exhibited an increase of the deflection of the galvanometer from approximately 40 to 120 mm, whereas the poisoned muscle under the same load — from approximately 50 to 240, that is, almost twofold greater. Consequently, in this respect too there exists, apparently, a correspondence between heat formation, or the intensity of resting metabolism, and the strength of the current of rest. But if the current of rest is conditioned by the intensity of resting metabolism, or by the intensity of the energy-supplying processes which maintain the muscle in its state, and if the role of stimulation consists precisely in the diminution or switching-off or disturbance of these processes, and consequently of this state, then of course the current of rest too must diminish during stimulation; such a phenomenon is indeed generally known under the name of “negative variation”. Thus, the contraction is the result not of an energy-supplying process, but, on the contrary, of the switching-off, or weakening, of such a process, and the external work takes place at the expense of the free energy of the system, of the change of the systemic conditions, consequently at the expense of the free energy of the structure. The raising of the intensity of the energy-supplying processes serves for the restoration of the systemic conditions, of the non-equilibrium state, and thus for the restoration of the capacity for work of the system, of its structural conditions.
That loading itself, without any contractions and stimulations, leads to an accelerated decomposition of creatine-phosphoric acid, I was able to show in certain unpublished experiments carried out together with my collaborator S. D. Borzdyko.
But if stimulation does indeed lead to contraction in such a way that, by means of a disturbance or switching-off of the energetic processes maintaining the structure, it disturbs this structure, then from this it follows directly that the more intensively these energy-supplying processes — conditioning the surplus of tension, and hence the non-equilibrium state — proceed, the stronger, consequently, must be the stimulating action, in order to retard the energy-supplying processes, that is, to disturb the structure and thereby provoke the process of equalisation of the system — the contraction. Thus, we arrive at the conclusion that the more intensive the resting metabolism, the higher must lie the threshold of stimulation. As was shown above, resting metabolism depends in a quite definite manner upon the load, increasing up to a definite limit, approximately corresponding to the maximal capacity for work of the given muscle. Consequently, the threshold of stimulation too must exhibit a similar dependence upon the load. To verify this conclusion we again took the gastrocnemius of the frog. The muscle was stimulated directly by a make-and-break induction current by means of two silver electrodes thrust into the upper third of the muscle. As the threshold of stimulation there was taken, as is customary, the maximal distance from the coil at which the first, scarcely noticeable contraction appears. In order to forestall the possible objection that the rise of the threshold upon loading is only apparent and is explained by the greater difficulty of observing the first contraction — since the load is greater and the possible isometric contraction sets in already earlier, but cannot be noticed — in order to forestall this objection, we again, as in the double myograph, clamped the muscle in the middle. In this way the lower half was mechanically stretched by the weight, while the electrodes were applied to the upper, unstretched half, on which the threshold of stimulation was determined. By this there was simultaneously excluded also the possibility that the change of threshold is determined in some way by those changes in the mechanical, elastic properties of the muscle which are directly provoked by the mechanical force of pull from the load. Proceeding from our assumption that at a greater load the muscle performs greater work and thus maintains the inequality of the tension of the muscle and the pull of the weight, we of course had to assume that the intensification of metabolism provoked by this would not be confined to the lower part of the muscle, and that, consequently, the change of the threshold of stimulation would tell also in the upper part, not tensed directly by the weight.
In the following table we adduce two typical experiments under loading by not too great weights (up to 50 g).
Table 1. Changes of the threshold of irritability under loading in the Gastrocnemius of the frog.
| Time (Exp. 8/I–1932) | Load | Threshold | Time (Exp. 9/I–1932) | Load | Threshold |
|---|---|---|---|---|---|
| 1 h | Unloaded | 295 | 5 h | Unloaded | 350 |
| 1 h 01 min | 40 g | 260 | 5 h 05 min | 50 g | 335 |
| 1 h 03 min | Weight removed | 290 | 5 h 06 min | 50 g | 330 |
| 1 h 05 min | 40 g again | 265 | 5 h 08 min | 50 g | 320 |
| 1 h 07 min | Weight removed | 275 | 5 h 09 min | 50 g | 315 |
| 5 h 10 min | Weight removed | 325 | |||
| 5 h 12 min | Weight removed | 330 | |||
| 5 h 14 min | Weight removed | 335 | |||
| 5 h 16 min | Weight removed | 340 |
The figures indicating the threshold signify, in millimetres, the distance between the primary and secondary coil in the apparatus of du Bois-Reymond.
The first experiment shows simply a rise of the threshold upon loading and a lowering of it upon removal of the weight. The second, however, is to show the course of the changes of that state which lies at the basis of the changes of threshold and is provoked by loading or by its removal. From this experiment we see that the change of state provoked by loading in the upper, unstretched part of the muscle develops and intensifies gradually over the course of 4–6 minutes. This is a further proof in favour of the view that we are here dealing not with a mechanical effect provoked by loading, but with a developing process of adaptation in the sense of excitation, that is, of an intensification of resting metabolism. If one represents graphically the course of this rise and fall of the thresholds after loading or removal of the weight, and compares the curve obtained with the course of the directly measured intensity of heat formation (for example, in Feng, on the sartorius of the frog after loading with various weights and after their removal — see Fig. 2 of the cited work), then we shall find a great similarity between the two curves: heat formation too increases strongly immediately after loading and then increases further over the course of 4–6 minutes.
We have shown, however, that the strength of the current of rest, and correspondingly of resting metabolism too, can increase with the load only up to a known limit, while at greater weights, exceeding this limit, it must again diminish. Therefore, if the threshold of stimulation depends upon the intensity of resting metabolism — in such a way that for the disturbance of the structure maintained by the energy-supplying processes a stimulation the more strong is necessary the more intensively these processes proceed, the higher the intensity of resting metabolism — then this threshold too must again diminish when a definite weight is exceeded. This indeed also takes place. The limit up to which the threshold still rises, and after which it already falls, is of course just as individual as for the current of rest or for heat formation, and fluctuates somewhat in dependence upon the state and power of the muscle. In the following table there is adduced a typical experiment in which this limit was attained at 150 g.
Table 2. Changes of the threshold of stimulation in the Gastrocnemius of the frog under loading by various weights.
| Time (Exp. 9/I–1932) | Load | Threshold (in mm) |
|---|---|---|
| 7 h | Unloaded | 250 |
| 7 h 02 min | 50 | — |
| 7 h 07 min | 150 | 235 |
| 7 h 14 min | 150 | 230 |
| 7 h 15 min | 150 | 240 |
| 7 h 17 min | 150 | 245 |
| 7 h 25 min | The whole weight removed | 250 |
| 7 h 23 min | The whole weight removed | 245 |
Thus, we see that our conclusion, derived from the assumption that the role of stimulation consists in the disturbance of the structure, or of the energetic processes of resting metabolism maintaining the non-equilibrium of the system — that this conclusion is fully confirmed in the experiment. Our theory permits us to predict the changes of the threshold of stimulation on the basis of the change of the intensity of metabolism or of the current of rest.
Let us mention further that, applying our general biological principle and the conclusions following from it to the physiology of muscles and to the investigation of the role of stimulation, we obtain a good explanation for a whole series of phenomena from the domain not only of the physiology of muscles, but of the physiology of stimulations in general.
With regard to muscles, let us adduce, for example, the fact that a muscle exhausted by stimulation does not restore its contractility if the stimulations continue, but, on the contrary, instead of this, falls into a state of prolonged contraction.[Author’s note:] Embden, S., Klin. Wschr., no. 29, 1930.
Here the role of stimulation as a brake, or as a disturbance of the energetic processes leading to the state of rest and maintaining it, is clearly seen.
In an exhausted muscle stimulation no longer provokes contractions, but retards the restoration which would set in in the case of the switching-off of stimulation, and leads to rigor, that is, to the equilibrium state setting in at death.
That the role of stimulation consists in the disturbance of the biological system in the sense of a disturbance of its non-equilibrium state, that is, of the energetic processes maintaining the structure, or in a direct damaging of the structure itself, whereby the conversion of the free energy of these processes into work against equilibrium is disturbed; that thus the reaction to stimulation is a process of equalisation, such as takes place at death too, with the sole difference that in the first case it is reversible, and in the second not — that all this is so, can be shown on all the phenomena of stimulation.
Let us recall certain phenomena which in the majority of manuals and textbooks are adduced as typical examples of stimulation.
Strong mechanical or chemical stimulations provoke a complete rounding-off of amoebae,[Author’s note:] E.g. M. Hartmann, Allgemeine Biologie, S. 131, Jena, 1925. and, as is known, when amoebae die, they too round themselves off, assume the minimal surface corresponding to equilibrium — the form of a sphere.
Zoothamnium alternans, Vorticella, as is known, contract upon stimulation; Koltsov,[Author’s note:] Biol. Zschr., Bd. III, H. 1 und 2, Moscow (in Russian), 1911. however, clearly observed that upon the dying of this form a death-contraction sets in, and upon fixation Zoothamnium is fixed in this state. Koltsov himself considers it beyond doubt that the cause of the death-contraction and of the contraction as a result of
stimulation is one and the same.
It is also known that the phenomena of stimulation in the mimosa are conditioned by a raising of permeability and an equalisation of concentration in the supporting cells, as takes place at death too.
One could adduce yet many such examples.
The same dependence between irritability, that is, the minimal magnitude of the impulse necessary to provoke a disturbance of the state of the structure and an approach to equilibrium, on the one hand, and the intensity of the basal metabolism, in other words, the distance from the equilibrium position, on the other, was able to be shown by V. A. Muzheev also on the smooth muscle m. retractor pharyngis of the snail (Helix pomatia). The state of the muscle, or the intensity of the metabolism maintaining this state, he influenced, on the one hand, by various, as far as possible constant, temperatures, and on the other — just as in the striated muscle — by various loads. A raising of temperature leads, generally speaking, in every tissue to an intensification of metabolism up to a known limit. Correspondingly to this, it was to be expected that with a raising of temperature up to a known limit the threshold of stimulation too would increase, in order thereafter, when a still higher temperature disturbs metabolism (the limit of adaptation of the tissue), to diminish again.
From this work[Author’s note:] Biolog. zhurnal, I, II, vyp. II–III, 1933. we adduce one table, which shows the dependence between temperature and the threshold of stimulation under a load of 2 g.
| Temperature (C°) | Threshold of stimulation* |
|---|---|
| 20 | 5 |
| 25 | 10 |
| 30 | 10 |
| 32 | 12 |
| 35 | 9 |
| 38 | 7 |
| 40 | Contraction |
| 25 | 6 |
[Author’s note:] *The distance of the contact of the potentiometer from the clamp of the positive pole, in centimetres. The voltage at the clamps of the accumulator is 1.5 volts.
We see that up to a known limit, in the given case up to 32°, the threshold increases, and then again diminishes. At 40° a spontaneous contraction sets in. Muzheev observed such spontaneous contractions in the majority of cases when the temperature passed beyond the limit of adaptation; this proves that when the temperature attains a magnitude damaging the muscle, irritability does indeed increase.
That temperature, and the change of the intensity of metabolism provoked by it, influences the state of the muscular tissue, is shown by the fact that upon a raising of temperature the muscle lengthened by 2–3 times, and then, upon cooling, shortened to its initial magnitude, although its weight did not change. The intensification of the processes of metabolism provoked by the higher temperature leads to a stretching, whereby it is proved that the energy-supplying processes of metabolism serve for the maintenance of the stretched state, and not for contraction and the production of external work. On the contrary, the shortening sets in when the intensity of metabolism falls in consequence of cooling, or when a disturbance is provoked by means of stimulation.
Analogous results were obtained by V. A. Muzheev also under various loads. But at a high temperature it was to be expected that the limit above which the weight itself would act harmfully and provoke a diminution of the threshold — that this limit lies lower, since the temperature itself already raises the intensity of metabolism; we had, consequently, to expect a summation of both factors, acting in one direction. Muzheev was able to confirm this conclusion too. From his experiments it is evident that a muscle with a load of 2 g exhibits spontaneous contractions at 40–41°, and with a load of 5 g — already at 32–33°; while under a load of 8 g the threshold diminishes as soon as the temperature becomes higher than room temperature (17°), and spontaneous contractions set in already at 22°.
Thus, all the adduced experiments show that stimulation provokes an impulse-conditioned disturbance of the non-equilibrium state of living matter, and that the energy-supplying processes of metabolism are needed not for the reaction, but, on the contrary, for the maintenance of the state of non-equilibrium, or for the restoration of it, which takes place without external influences after each response to stimulation. Thus, after each disturbance provoked by an impulse, the laws of motion of living matter again come into their own. Consequently, external work in the physical sense can be produced directly only at the expense of the structural forces themselves, during and in consequence of the disturbance of the structure.
In recent years more and more facts confirming this point of view have been accumulating, and the number of biologists is increasing who consider that the energy-supplying processes are needed precisely for restoration, and not for the contraction of the muscle. The same is sought to be proved, for example, by Ritchie too.[Author’s note:] Ritchie, Theories of muscular contraction, The Journal of Physiology, vol. 78, 1933. But here it is important to emphasise that the matter concerns not only the necessity of the energy-supplying processes for the restoration of the state of rest, in so far as the latter possesses a greater potential energy, but also the fact that this state of rest is a non-equilibrium one, that is, that the energy-supplying processes are needed not only for its restoration, but also for its preservation, since only in this way does the role of stimulation, as a disturbance of the laws of motion of living matter as a result of an external impulse, become clear to us. It should be noted that, specifically in respect of the muscle, this assumption was put forward by Gurwitsch,[Author’s note:] Gurwitsch, Die histologischen Grundlagen der Biologie, Verl. Fischer, Jena, S. 224, 1930. who, analysing the functional changes of the structure of striated muscle, arrived at the conclusion that “one must take into account the possibility that a fully exhausted (or narcotised) muscle is in a state of its elastic equilibrium, while a normal, resting, tonic muscle, on the contrary, is dynamically stretched beyond its equilibrium. True contractures would then be the correlate of a complete or almost complete cessation of known dynamic processes in the muscle, while the whole scale of active states of the muscle (including relaxation), on the contrary, would be the visible expression of definite forms of its activity.”
It remains for us to show that stimulation does indeed provoke a disturbance of the non-equilibrium structure of living matter, and that the energy-supplying processes are indeed needed for the restoration of this structure.
As we have already said in the chapter on the structure of living matter, its molecules, or the micelles formed from them, are in a stretched, deformed state, and possess in consequence of this an elongated form and a raised dipole moment, and by virtue of this a raised tendency to association. There too we have already spoken of the experiments of K. Meyer,[Author’s note:] Biol. Zschr., Bd. 214, 1929. who established by means of X-ray photographs the shortening of the micelles upon contraction of the muscle. His calculations, made on the basis of these X-ray photographs, showed that the maximal contraction may amount to 80%, which agrees well with the data obtained directly on the muscle. The contraction, and consequently the mechanical work upon the lifting of a weight, takes place at the expense of a change of the molecular structure, as is required by our principle of the work of structural forces. Another proof in favour of the disturbance of the state of living matter produced by stimulation is the already-mentioned negative wave, and the fact that at the moment of contraction the optical anisotropy of the muscular tissue diminishes. In striated muscle this is expressed in a lesser clarity of the transverse striation, in a more or less pronounced homogenisation. Such a reversible homogenisation in living muscles of Hydrophilus was described already by Hürthle[Author’s note:] Hürthle, Pflüg. Arch. ges. Physiol., Bd. 126, 1909. in 1909. More recently B. Holz[Author’s note:] B. Holz, Pflüg. Arch., Bd. 230, 1932. established by means of more exact methods on the living m. cutaneus pat. of the frog that the ratio of the number of anisotropic bands to the number of simply refracting ones changes upon contraction in favour of the latter. The muscle taken by Holz is very convenient for such observations, since it consists almost exclusively of a single layer of parallel fibres. Holz mentions that upon direct stimulation of the muscle
homogenisation sets in, and after the cessation of stimulation the transverse striation again emerges, sometimes still more sharply than before. But since anisotropy is based upon the elongated form and the parallel arrangement of the micelles forming the fibres, that is, since it is a double refraction on the basis of form, the diminution of the double refraction serves as the expression of the fact that the micelles have shortened and approached equilibrium, or, at the least, that their parallel arrangement has been disturbed. However, in the chapter on the structure of living matter we saw that both these factors are not independent of one another. The parallel arrangement in the fibres is provoked precisely by the association-tendency of the dipoles; therefore when the micelles shorten and their dipole moment diminishes, the tendency to association too is lowered, which, in the end, leads to the disappearance of the fibrous structure.
Upon dying, when the energetic processes conditioning the stretched state of the molecules cease, the transverse striation and the fibrous structure must consequently disappear. In order to prove that the energy-supplying processes do indeed go to the maintenance of this structure of the molecules of living matter, we must show that the duration and degree of the double refraction, or of the fibrous structure, stand in direct dependence precisely upon those energetic processes by which the preservation both of the stretched state at rest and of the current of rest is conditioned. Investigations of these relations we carried out together with V. A. Muzheev.[Author’s note:] Biol. zhurl., t. III, vyp. II–III, 1933.
The object served us by the m. cutaneus pectoris, which is so thin that it can be investigated and photographed under the microscope while alive. The muscle was placed in Ringer’s solution, and from time to time we observed or photographed it. To illustrate the results of the experiment we adduce several figures. In Fig. 21 we see the gradual disappearance of the transverse striation and finally the disintegration of the fibres into granules.
The differences in the change of structure in the loaded and the unloaded muscle are shown by Figs. 22 and 23. The muscles were fixed under the microscope, one in the stretched, the other in the unstretched state, and from time to time exactly one and the same place was always photographed. Just as under the influence of stretching the creatine-phosphoric acid decomposes sooner, and correspondingly to this, after the initial rise, the current of rest in the end falls sooner, so here too in the stretched muscle we observe at the beginning a clear transverse striation, which, however, disappears considerably sooner. While in the unstretched muscle a noticeable disappearance of the transverse striation begins only after 4 hours, in the stretched muscle it sets in already after 2 hours, and after 4 the fibrous structure too has already disappeared, and we see almost wholly granular tissue. This result is fully comprehensible in view of the small size of the muscle and the inconsiderable reserves of energy connected with this.
Figs. 24 and 25 show the changes after prolonged stimulation, that is, upon fatigue and after rest, in a normal muscle and in one poisoned with monoiodoacetic acid. We see that a known homogenisation, or a diminution of the clarity of the transverse striation, sets in in the normal muscle immediately after prolonged stimulation, but this striation again emerges after a 20-minute rest. In the poisoned muscle, however, where the resynthesis of phosphagen is impossible and the reserve is rapidly exhausted, the transverse striation disappears finally.
If, finally, we take a poisoned muscle and place it in the stretched state under the microscope in Ringer’s solution, then the transverse striation, at first very clear, disappears already after 70–80 minutes, as is seen in Fig. 26, since stretching provokes an accelerated decomposition of creatine-phosphoric acid, and since the latter, in consequence of the poisoning, cannot be synthesised anew, its reserve is rapidly exhausted, and the source of energy for the maintenance of the transverse striation and the fibrous structure disappears.
Thus these experiments graphically show that stimulation does indeed provoke a disturbance of the non-equilibrium state of living matter, and that the energy-supplying processes are indeed needed for the maintenance or restoration of this structural state.






Finally, we shall endeavour to represent to ourselves more clearly in what the disturbance provoked by stimulation properly consists. According to our theory of living matter, the molecules of the latter are in a deformed, excited state. They are maintained in this state by means of metabolism. For this, as has already been said, it is necessary that the molecules of living matter should collide with molecules before they have come into the equilibrium state, that is, a constant process of assimilation must be secured. For individual gas molecules in rarefied gases, where the van der Waals forces of attraction may be neglected, the “length of life” of an excited molecule equals, as is known, 10⁻⁸–10⁻⁷ seconds. If, however, the molecules are associated, or, the more so, included in the lattice of a crystal, so that one can no longer speak of individual molecules, then the equalisation of the excited deformed state will last considerably longer. In dependence upon the mean length of life of a deformed, excited molecule of living matter and upon the mean duration of the reaction with a food molecule, that is, of the process E″ + N → E′N′ + n₁ + n₂ + … + Q → E′, the structure must, for the unhindered maintenance of the non-equilibrium state, secure a known mean frequency of collisions with food molecules. In other words, a known intensity of metabolism or of assimilation must be secured. When, however, an impulse provokes a disturbance of the structure, by virtue of which the probability of collision diminishes, then the non-equilibrium state is disturbed, and the molecules will begin to approach equilibrium. Such a disturbance may consist in a lowering of the associative forces owing to concussion or to electrical forces. On the other hand, the lowering of the intensity of metabolism in consequence of the approach to equilibrium and the diminution of the dipole moment leads to a weakening of association. After the cessation of the disturbing action of stimulation, unhindered assimilation again sets in, and the structure is restored. By this one can without difficulty explain the phenomenon described by Muralt[Author’s note:] Muralt, Pflüg. Arch., Bd. 230, 1932. in the investigation of the changes of double refraction in the muscular tissue during contraction. Muralt investigated not the change of the transverse striation, that is, the ratio of the width of the anisotropic bands to the width of the isotropic ones, but the degree of double refraction of the whole muscular tissue, registering photographically the displacement of the interference bands of two rays passing through the tissue. It turned out that during contraction there does indeed take place a diminution of double refraction. Besides this, it turned out that between the diminution of double refraction during contraction and the subsequent intensification of it upon relaxation there is observed yet a small wave, so that one can clearly distinguish four phases: a pronounced diminution of double refraction, a slight intensification of it, a subsequent diminution, and, finally, an intensification to the initial magnitude. It is accepted by all investigators, and this stands in accordance with our theory of living matter, that the double refraction is a refraction based upon the elongated form and the parallel arrangement of the micelles in the fibres. The diminution of double refraction is consequently conditioned in the first place by the fact that stimulation provokes a disturbance in the direction of a shortening of the micelles and a weakening of the associative forces. Thereby, however, the micelles approach the equilibrium state, that is, the undeformed lattice structure, which for its part conditions an intensification of the so-called intrinsic double refraction (see Chapter IV). By this the second phase is explained. But now unhindered assimilation, restoration, again sets in. The molecules enter into reaction and are again stretched, deformed, at the expense of the energy liberated upon the decomposition of the food molecules; the inconsiderable intrinsic double refraction disappears (third phase). Thereby the dipole moments, and consequently the associative forces, are again intensified, the micelles arrange themselves in parallel, and the original double refraction of form[Author’s note:] Formdoppelbrechung of the German authors. is restored (fourth phase). The shifts, noticed by Muralt upon fatigue, of the relations of these phases in their temporal succession in the direction of a relative lengthening of the third phase (that is, of the restoration of the non-equilibrium structure and the degree of deformation) are likewise fully explained in this way.
Since we have assumed that the raising of the potential of the molecules cannot take place as a result of the onset of metabolism after its disturbance, but that, on the contrary, this potential would have to fall still further, the stretching of the molecules cannot be explained by their assimilating food molecules themselves. The energy necessary for the stretching they must receive at the expense of the energy of other living molecules of the tissue, and these latter must in consequence of this exhibit a continuous fall of potential, in so far as the fundamental process does not set in. Such a continuous fall of potential is expressed in fatigue. The degree of stretching setting in after each contraction becomes ever smaller. If after fatigue a prolonged rest is interposed, then restoration sets in. In consequence of the strong lowering of the potential of the tissue during this rest, the fundamental process sets in, the potential again rises, the molecules of the working structure can again be stretched to the former degree. In the preceding chapter we saw that the fundamental process is accompanied by an intensification of respiration. In just the same way we find in the muscle too that restoration can take place only in the presence of oxygen. By this one could explain also the hypertrophy of a muscle provoked by exercise, that is, the increase of the mass of the muscle, since by the raising of the potential the possibility of assimilation with an increase of mass is given.
Chapter 4: Evolution
In the previous chapter we reached the conclusion that if certain magnitudes of the state of a definite environment exceed a known limit, then the action of this environment leads to a disturbance of the process of metabolism and, in consequence, to an injury of living matter. In such cases we said that the boundary of physiological adaptation had been exceeded. But, as we likewise indicated, the injury of living matter arising in this way may lead to the fundamental process. In that case the fundamental process is called forth by the action of the external environment and, accordingly, sets in earlier than it would have set in within the boundaries of physiological adaptation. Here the expression “earlier” is to be understood not in the sense of time, but in the sense that the fundamental process sets in even when the free energy has not fallen to that degree at which it sets in if metabolism is not disturbed. The fundamental process itself, however, we excluded from the number of phenomena of physiological adaptation, since, although it is indeed called forth by an external action and secures the preservation of living matter under these altered conditions, it nevertheless is precisely an indication that the boundaries of physiological adaptation have been exceeded. We had all the more right to proceed thus inasmuch as the life cycle of a living system, an organism, is determined, as we have seen, by the laws of metabolism, whereas it is upon the fundamental process that reproduction is based, that is, the formation of new individuals, or of a new generation. The same applies also to regeneration: here too it is no longer the former cells or tissues of the individual that adapt themselves to the altered environment, but a new cellular generation, arisen out of the embryonic cells (see the chapter “Reproduction”). Here, consequently, we can no longer speak of the physiological adaptation of any organism, but only of the adaptation of living matter in general. But to this too we have a right, of course, only in the case where the fundamental process does indeed lead to the formation of such new organisms for which this altered environment, which called forth the premature fundamental process, still lies within the bounds of physiological adaptation. In this case there must arise, as a result of the fundamental process, germ cells with other properties, with another structure. For, as we showed in the previous chapter, the boundary of physiological adaptedness, the criterion of which was the preservation of Rubner’s constant, depends exclusively upon the special hereditary structures of the germ cells and upon the magnitudes µ₀ and c characterising them. But, as is known, the special hereditary structures are determined by the fine structure of the chromosomes, and by this same fine structure the magnitude c too is probably essentially determined, whereas Rubner’s constant, it would appear, depends in the main upon µ₀. This means the following: if we have two organisms adapted to one and the same environment, that is, in which, by virtue of their hereditary structures (which need not be identical), no disturbances of the laws of metabolism are observed in the given environment, so that the quantity of energy liberated by them over the whole of their life per unit of mass corresponds to the magnitude of their Rubner’s constant — if we have two such organisms, then the value of Rubner’s constant is determined in the main by the magnitudes µ₀, m₀, that is, by the free energy of the germ cells. Therefore organisms with a very different hereditary structure may possess one and the same Rubner’s constant and, conversely, organisms with a very different constant may possess one and the same structure, that is, be adapted to one and the same environment. Thus, for example, terrestrial and aquatic animals may possess eggs with the same free energy and, by virtue of this, the same Rubner’s constant, although the fine structure of their chromosomes is quite different, in consequence of which one of the animals has organs and characters making it suited for life on land, and the other — for life in the water. Conversely, two animals
furnished with gills may form eggs with very different free energy and, in consequence of this, possess different Rubner’s constants too. Thus, the formation of germ cells with a new structure is a necessary precondition for the adaptation of living matter beyond the boundaries of physiological adaptability. But, as is known, every organism always forms germ cells with one and the same fine structure, whereby the preservation of the specific properties from one generation to another is secured. This preservation of the fine structure of the chromosomes and thus of the specific properties, that is, the phenomena of heredity [наследственность — nasledstvennost′], and their laws discovered by Mendel, are secured by the mechanism of cell division, of fertilisation, and of the maturation divisions in the sexual cells. The formation of germ cells with a new structure, that is, in the language of genetics — mutation, has as its precondition a disturbance of this mechanism, which serves for the preservation of the fine structure in the formation of germ cells. But since the formation of full-value mature germ cells is based upon the fundamental process (see the chapter “Reproduction”), it is natural to assume that the adaptation of living matter beyond the bounds of physiological adaptability, that is, mutations, is based upon a disturbance of the fundamental process. Disturbances of this process are to be expected, above all, in the case where it sets in not at the time which would correspond to undisturbed metabolism, but “prematurely”, in consequence of a disturbance of the latter. But this takes place precisely when we pass beyond the boundaries of physiological adaptation. If, then, to physiological adaptation in the sense of a disturbance of the laws of metabolism we oppose the adaptation of living matter in the sense of the formation of new hereditary structures, calling it for brevity “genetic adaptation” [генетическое приспособление — geneticheskoe prisposoblenie], then we may formulate our conclusion in the following manner: when known magnitudes of the state of the external environment pass beyond the boundaries of physiological adaptation, then, as a result of the injury of living matter and of the disturbance of the fundamental process occasioned thereby, genetic adaptation sets in; or, more briefly: there, where physiological adaptation ceases, genetic adaptation comes into force. The expression “genetic adaptation” should not give rise to misunderstandings: it here signifies nothing other than the appearance of hereditary changes, that is, of mutations, and says nothing about the kind or the direction of these changes. Since we are dealing with a disturbance of the normal course of the processes serving for the preservation of the fine structure of the chromosomes — the processes securing heredity — there can be no question of the arising of mutations conditioning such properties by virtue of which the organism is made adapted to a new environment; what is in question here is only that mutations arise in general, and that in this way the possibility is given of the appearance of adapted mutants as well.
Before occupying ourselves with the consequences which this connection between the boundary of physiological adaptation and the arising of hereditary changes has for evolutionary doctrine, we shall investigate to what extent our supposition, made on the basis of our general laws and of the considerations set forth above, is confirmed by experimental material. If the changes of the structure of the hereditary mass conditioning mutations are based upon disturbances of the formation of full-value germ cells, that is, upon disturbances of the fundamental process, then it is natural to expect that 1) mutations occur precisely during the fundamental process, that is, during the formation of mature germ cells; 2) that mutations are called forth by the injury of living matter and the consequent raising of the charge of the germ cells; and finally 3) that the injury of living matter leading to the fundamental process does indeed call forth a disturbance of the processes of formation precisely of the hereditary mass, that is, of the nuclear apparatus of the chromosomes and their structure.
As regards the first point, Muller,[Author’s note:] Muller, Genetics, July, p. 279, 1928. in his investigations of the action of temperature upon the quantity of mutations, established that, in respect of the acceleration of the mutational process, it is chiefly the period of maturation — that is, of the formation of full-value sexual cells — that is sensitive to temperature. Muller himself also discusses the possibility that the action calling forth mutations, in the given case temperature, acts not upon the ready-made, pre-existing fine structure, but disturbs the process of formation of new genic
material, that is, that the “mutation” consists in this, that for some reason the genic material newly formed during the period of action is not identical with the former. According to Muller, the elective action of temperature upon the speed of the mutational process precisely during the period of maturation would be explained by the fact that temperature itself accelerates the process of growth of the cells and thereby of the formation of new genic material, that is, that precisely during the action of the high temperature, which conditions the disturbance of the new-formation, more genic material is formed. Consequently, according to this conception, high temperature ought not to call forth mutations when acting upon already matured sexual cells.
As regards the second point, all the data of experimental genetics speak in favour of the fact that mutations are called forth by the injury of living matter, that is, by such actions as lie beyond the bounds of physiological adaptation. Thus, we know that the actions accelerating the mutational process, such as Röntgen rays and high temperature, call forth a noticeable effect only in the case where they are applied in an injurious dose. In exactly the same way, according to Hanson and Heys,[Author’s note:] Hanson, F. B. and Heys, F., American Naturalist, vol. 67, 1933. the action of the rays of radium, it would appear, is heightened if the organism is narcotised, that is, injured.
That the injury need not necessarily affect the living matter of the germ cells themselves (inasmuch as, as was shown in the chapter on reproduction, the fundamental process in multicellular organisms is not limited to a single cell), was proved by the experiments of Gülbenkian,[Author’s note:] Gülbenkian, Biol. zh., t. III, no. 2, 1934. undertaken in part on my initiative. Gülbenkian acted with identical doses of Röntgen rays upon the whole body of the fly Drosophila melanogaster, and separately upon the rear part alone, containing the sexual cells, and then upon the front part alone, not containing them; the results thus obtained in respect of the frequency of mutations were compared with two control series, of which one served to verify whether the action of secondary or scattered rays might have taken place here. The results obtained were as follows:
| Category of flies | Number | Number of lethal mutations | % of lethal mutations |
|---|---|---|---|
| Whole body irradiated | 642 | 28 | 4.36 ± 0.8 |
| » rear end | 925 | 18 | 1.94 ± 0.41 |
| » front end | 1 140 | 16 | 1.39 ± 0.35 |
| Control inside the chamber | 971 | 1 | 0.10 ± 0.15 |
| » ordinary | 514 | – |
A statistically significant difference is thus observed everywhere except in the difference between the two control series. The difference, however, between the flies in which the whole body was irradiated and those in which only the rear end was irradiated exceeds the mean error by 2.63, that is, almost threefold.
These experiments show clearly that, for the acceleration of the rate of mutation, there is no need of a direct action of the rays upon the germ cells: it suffices to irradiate only the body without these cells. The experiments admit of no other explanation. Were we to adhere strictly to the figures, we should have to arrive at the further conclusion that what is decisive is the magnitude of the injured portion of the body, and not whether germ cells are contained in it or not, since between the second and third series (irradiation of the front and rear ends) there is no real difference: the latter exceeds the mean error by only 0.36.
Finally, as regards the third point, according to which the injury of living matter leading to the fundamental process calls forth a disturbance of the processes of formation precisely of the hereditary mass, that is, of the nucleus and the chromosomal apparatus, in order to prove this assertion we must digress somewhat further. The point is that here there is observed not a particular dependence between these two processes, playing a role only in the appearance of mutations. On the contrary, we are here dealing with a general dependence, of which the appearance of mutations is merely a particular case.
In order to prove the presence of such a general dependence between the injury of living matter and the disturbance of the structure of the nucleus, we shall start from the representations and experiments set forth in the chapter on reproduction. There we showed that between metabolism and the fundamental process there exists an inverse dependence, and that reproduction, that is, the fundamental process upon which it is based, sets in precisely when metabolism in the sense of the assimilation of non-living molecules can no longer take place. But reproduction itself is essentially connected with the distribution of the hereditary mass among the newly formed individuals, that is, with the formation and alteration of the structure of the nuclear apparatus. Proceeding from these considerations, we were able to show that the reconstruction of the nuclear apparatus in Oxytricha takes place only when the inflicted injury does indeed lead to the fundamental process, that is, in ageing individuals, and that this reconstruction does in fact set in regularly if the injury leads to the fundamental process; and even in normal division — which is also, after all, conditioned by the gradual weakening of metabolism and by the fundamental process setting in in consequence thereof — we likewise have before us a mobilisation, that is, a reaction of the nuclear apparatus. The very same mobilisation of the nuclear apparatus, by means of an artificial injury of the cell, we observe also in germ cells in the well-known experiments with artificial parthenogenesis. Here one can compel the nucleus to divide and call forth development by means of any non-specific injury of the cell. But this connection exists not only in unicellular organisms and eggs, but also in highly differentiated cells of multicellular organisms. Thus, for example, in the so-called atrophy in consequence of inactivity, in striated muscle there is observed a thinning and resorption of the muscle fibres, accompanied by a considerable increase in the number of nuclei. The same is observed also in nerve cells upon the severance of the nerve processes and the degeneration beginning in consequence thereof. That this mobilisation of the nuclear apparatus in multicellular organisms is not limited to the nucleus of the injured cell itself is shown by the facts adduced in the chapter on reproduction. Thus it becomes, at the least, highly probable that this process of reconstruction of the nucleus and of the formation of new nuclear material may be disturbed more easily if it takes place “prematurely”, that is, not when it is called forth physiologically as a result of the extinction of metabolism. This is probable for two reasons: in the first place, the nucleus possesses both its own metabolism and its own changes of state, proceeding within the framework of the cycle of the cell or of the organism. Therefore, if an external injury calls forth the fundamental process earlier, then the nucleus too is mobilised earlier; consequently, a forced, hurried reconstruction and distribution of the hereditary mass sets in. In the second place, in this case the dissimilatory processes conditioning the fundamental process are more intense. The crudest expression of the disturbance, setting in, of the reconstruction and distribution of the hereditary mass is this, that whereas in the physiological onset of the fundamental process it leads to reproduction and, in consequence, to a uniform distribution of the hereditary mass, in such cases there often takes place no uniform distribution of the hereditary mass at all, and in consequence no division takes place either, as for example in our experiments with Oxytricha. The same thing, that is, the reconstruction of the nuclear apparatus without normal division, takes place in unicellular organisms in consequence of unfavourable conditions and in the formation of cysts or in endomixis. Jennings with his collaborators[Author’s note:] Jennings, H. S., Raffel, D., Lynch, R. S. and Sonneborn, T. M., J. of exp. Zool., vol. 62, p. 363–408, 1932; Caldwell, L., ibid., vol. 66, no. 3, 1933. were indeed able to prove the appearance of new hereditary forms after endomixis. True, it cannot be proved that here a true mutation of one or several genes does in fact take place, since experiments with crossing are here impossible. The supposition is admissible that in these cases we are dealing with the splitting-out and manifestation of recessive characters. But this is of no significance for the fact that here the distribution or reconstruction of the hereditary mass in any case takes place otherwise than in ordinary division, that is, that this process is disturbed. For indeed it is by no means obligatory that every disturbance of the fundamental process or of the reconstruction of the nuclear apparatus connected with it should consist in a genic mutation. What is in question is only that a genic mutation too is merely a special kind of this disturbance. The latter may
manifest itself in the dropping-out of division, in polyploidy, in irregular splitting-out or chromosomal aberrations, and finally also in a disturbance of only the finest structures, that is, in genic mutations. Here, again, it is probable that certain of these kinds of disturbance occur in combination with one another.
One of the best proofs of the fact that a non-physiological raising of the dissimilatory processes, or injury in consequence of too strong a decomposition without assimilation (in the strict sense of the word), leads to a disturbance of the distribution of the hereditary mass and of its structure, is provided by the recently published experiments of M. S. Nawaschin.[Author’s note:] Nawaschin, Altern der Samen als Ursache der Chromosomenmutationen, Planta, Arch. f. Wissenschaftliche Botanik, Bd. 20, H. 2, 1933. The result of these experiments is that “in resting and ageing seeds there take place processes leading to a considerable increase in the frequency of chromosomal mutations, just as is observed under the influence of large doses of Röntgen rays”. Here, it would appear, it is beyond doubt that we are dealing with a non-physiological raising of the dissimilatory processes, that is, with injury. “In point of fact”, writes Nawaschin, “it is only in the hands of men that seeds can lie for years without germinating; in nature this occurs considerably more rarely, and the overwhelming majority of plants are adapted to a shorter period of rest in the seeds”. Thus, in accordance with our representations, we are here dealing with injury in consequence of the exceeding of the boundary of physiological adaptation. Nawaschin’s conclusion that mutations are in general founded upon the injury of living matter in consequence of a disturbance of the processes of metabolism likewise corresponds to our representations. He writes: “We must, then, concentrate our attention upon the totality of the diverse processes taking place within the living cell, instead of fruitlessly seeking ‘specific’ external exciters of the process of mutation in nature. Such external exciters, applied by us with the aim of artificially obtaining mutations, for example by means of rays, poisons, narcotics and the like, perhaps only alter the course of the vital processes, creating conditions similar to those which must have been created in a living cell which has been for a long time in an anabiotic state”. That these injuries, in the end, lead to a disturbance of the structure of the nuclear apparatus is incontestably evident from Nawaschin’s work. He describes various irregularities of the structure of the nucleus, arisen as a result of a disturbance of the processes of division and new-formation of the nucleus, for example numerous nuclear fragments lying in the protoplasm, cases of the agglutination of whole chromosomes, and so forth.
Thus, the data of experimental genetics, as we see, speak clearly enough in favour of the fact that the appearance of mutations is based upon a disturbance of the fundamental process, this disturbance being called forth by the passing of the magnitudes of the state of the environment (or of the duration of their action) beyond the boundary of physiological adaptation, which leads to the injury of living matter and to the onset of the fundamental process accompanied by a disturbance of the reconstruction of the hereditary mass. We see also — and this circumstance seems to us especially important — that this dependence is not only able to explain the immediate factual material of genetics, but itself finds its grounding in more general biological regularities, thereby permitting us to represent the regularities of mutations as particular cases of these general biological laws.
In considering the conclusions which follow from this connection between the boundaries of physiological adaptation and hereditary variability for evolution (that is, for the adaptation and development of living matter beyond the limits of individual existence), we arrive at a very simple representation concerning the significance of the surrounding environment, of the boundary of physiological adaptation and its criterion — Rubner’s constant — and also of mutations and of heredity itself for the formation of species. From what has been said it follows that the physiological adaptation of organisms to the surrounding environment and to its changes has no significance for the adaptation of living matter beyond the limits of individual existence, that is, for evolution and the formation of species, which sharply contradicts Lamarckism. Precisely so long as organisms are able to adapt themselves to the environment during their life, that is, so long as the fluctuations of the external conditions do not exceed the boundaries of physiological adaptation — precisely so long are the processes of metabolism not disturbed, Rubner’s constant is preserved, and there take place no injuries capable of leading to a disturbance of the fundamental process and thus to mutations. On the contrary, if in the surrounding environment there take place changes going beyond the boundaries of physiological adaptation, then, in consequence of the injury, mutations will arise. Consequently, in a new environment to which the given organisms are no longer adapted, there will be observed in them more frequent mutations, with the formation of germ cells differing from one another in their fine structure and in their initial potential, and, as a result, there will increase the probability that among the new variants which have appeared there will be found such as are adapted to the new environment too. If, then, the surrounding environment does not exert a direct influence upon evolution, this influence of it nevertheless makes itself felt in an indirect way, when the external conditions go beyond the limits of physiological adaptation. This influence is indirect inasmuch as we are here dealing not with the arising of mutations through the direct influence of the surrounding environment upon the structure of the hereditary mass, but with the manifestation of the laws of motion of living matter under these altered external conditions. This thought may also be expressed thus: the survival of adapted organisms in the struggle for existence is a conservative, species-preserving factor. On the contrary, non-survival in the struggle for existence is the moving, species-forming factor of evolution. The material for evolution is supplied not by the victors in the struggle for existence, but by the vanquished. All the organisms that have been preserved were, at their arising, adapted to the surrounding environment, for otherwise, after all, they could not have been preserved. The great thought of Darwin consisted precisely in this, that from this it by no means follows that only adapted organisms arise. There always arise, in large quantity, less adapted (or wholly unadapted) organisms as well, while the preservation of the adapted ones is the result of selection. But between the arising of the various variants and the process of selection neither Darwin nor contemporary biology sees any connection. The arising of hereditary changes is, in their opinion, a process given once and for all and independent of the process of selection and survival. Our considerations, however, lead us to the conclusion that here there is a causal dependence, representing a consequence of the general laws of motion of living matter. Precisely in so far as, and because, not only adapted organisms arise, in so far and for that reason do new hereditary variants appear.
In the assumption of the independence of the arising of hereditary variants from the physiological fate of organisms, it is not difficult to discern the same thought of the separation of soma and germ plasm which Weismann postulated in his doctrine of potential immortality. As there it is assumed that the germ plasm possesses its own laws and does not age, so here too it is assumed that the changes of the germ plasm proceed independently of what is taking place in the rest of the body. This independence of the two groups of processes is adhered to so strictly that contemporary geneticists are inclined rather to admit a direct action of the environment upon the hereditary mass than to allow an action of the body itself. This shows, as it seems to us, that if one does not reckon with the general laws of motion peculiar to living matter, as they are presented in the present book, then the most essential biological dependence and regularity of phenomena escape a more profound analysis.
In appraising the significance which this dependence between the boundaries of physiological adaptation and mutation has for evolution and the formation of species, it should be taken into account that we need not here necessarily be dealing with sharp changes of the surrounding environment, lethal for all the individuals of one species or even for the majority of the species of a large systematic group, that is, such as these species can no longer adapt themselves to; there may also occur insignificant or narrowly local fluctuations, exceeding the boundary of physiological adaptation only for separate individuals. Then precisely these individuals will give the new variants. With the first case we are probably dealing when one or several species die out, to be succeeded by a great diversity of other forms, as is known, for example, with respect to the dying-out of the large reptiles at the end of the Mesozoic and their replacement by the mammals in the Cenozoic.
All these considerations, together with the briefly set forth theory of the arising of mutations, concern exclusively certain regularities of species-formation, the conditions of the arising of new forms, but give no theory of evolution in the sense of a historical regularity, as we defined it in Chapter III of the general part. The question arises: is it possible, on the basis of our general laws of motion and of the historical principle of the increase of external work which we have given, to work out a more special theory of evolution, or at least to find certain approaches to it?
Proceeding from the theory of species-formation given in this chapter, we must seek the regularity in the properties of the new germ cells appearing upon the disturbance of the fundamental process. Such a regularity may consist in a regular alteration either of the special hereditary structures or of the magnitudes of the germ cells possessing a general significance, that is, of the magnitudes µ₀ m₀.
But the regularity of the alteration of the structure of the germ cells cannot be grounded in the regularities of the processes of formation of these cells themselves. The process of formation contains only the possibility of an alteration in consequence of an injury of the organism, whereas what is historically regular in these alterations can be given only in the case where the conditioning injurious factors do not themselves lie outside the system and are not independent of the laws of motion of the system under consideration. This is an essential property of historical regularity, as was shown in Chapter III. In application to our case of the formation of germ cells with an altered structure, this signifies the following: since the alterations of the structure of the germ cells, according to our theory of mutations and species-formation, are conditioned by the injurious actions of the surrounding environment, the cause of the historical regularity of the alteration of these structures can lie only in the laws of motion of such a system as also contains the surrounding environment acting upon the organism. And here the changes of the environment conditioning such a historical regularity must essentially depend upon the laws of motion of living matter. But the changes of the external environment acting upon organisms we may divide into two classes: changes of non-living and changes of living nature. The first class, that is, the changes of non-living nature, such as for example changes of temperature, of the composition of the air, of sunlight, the geological changes of the earth’s crust in the course of the development of the earth, and so forth, do not essentially depend upon the laws of motion of living matter. Were we to consider that for the historical regularity of evolution which we have deduced these changes too play a role, possessing in respect of it a fortuitous character, then we should have to regard the earth as a whole as a system with its own laws of motion, and either deny our historical regularity of the evolution of living matter or deduce it from such general laws of motion of the earth as a whole. Our assertion that the changes of non-living nature are independent of the laws of motion of living matter signifies, consequently, that living matter on the earth has not essentially altered the geological and meteorological history of the latter, and — what is more important for us — that the geological history of the earth has not essentially influenced the historical regularity, indicated by us, of biological history; the direction of geological history would have remained unaltered even if living matter had not chanced to arise on the earth, while on the other hand the regularity of the history of living beings, the general direction of evolution indicated by us, would have remained the same even if the earth had chanced to have another geological history too. Our assertion of course by no means signifies that the changes of non-living nature have not influenced the course and the paths of this evolution, that is, the paths of the realisation of this direction of development, or that under the influence of non-living nature other historical regularities have not arisen.
As regards the changes of the surrounding environment belonging to the second class, that is, the changes of living nature, these do essentially depend upon the laws of motion of living matter. As an injurious change of the living surrounding environment there must in the first place, of course, be adduced the density of population, depending upon the reproduction of the organisms themselves. The laws of motion of living matter contain, consequently, conditions which of necessity lead to its injury and thus to the formation of new forms. Thus, according to what has been said before, for what is historically regular in evolution (if it exists) significance can be possessed only by those injuries which are called forth by the living surrounding environment, or only by those new forms which secure survival, that is, victory in the struggle for existence under the altered living surrounding environment. Those new forms, however, which secure survival under a change of the non-living environment, are not essential in respect of what is regular in evolution. Moreover, from what has been said it follows that the historical regularity of evolution is a regularity not of separate organisms or even of species and their interactions with one another, but a regularity of the whole of living matter existing on the Earth.
This division in respect of the significance which the actions of living and of non-living nature have for the formation of new forms corresponds to that division at which we arrived in the chapter on the contradiction between external and internal work. There we said that there may arise new structures which permit either the utilisation of new sources of energy or the production of more work using the same sources of energy, and that, given identical structures or the utilisation of identical sources of energy, in consequence of reproduction and overpopulation the advantage will be possessed by those organisms which produce more external work, that is, that new forms may be preserved either because they utilise new sources of energy or, using one and the same sources of energy, because they are able to produce more external work. But adaptation to the injurious action of a change of the surrounding environment by means of the formation of new forms adapted to the new environment may, in the case of changes of non-living nature, take place in both directions. For example, with a change of temperature, say with a cooling, there may be formed new structures protecting against the giving-off of heat (fur and the like), or permitting the production of more intensive external work, and so forth, adapting in this way new forms to this change of the environment; whereas in the case of changes of living nature in consequence of overpopulation, the new forms called forth by such injury can be adapted to this injury only in so far as they produce more intensive external work. Of course, they may survive even without more intensive external work if they are better adapted to non-living nature, for example utilise other sources of energy of non-living nature, and so forth. But this does not signify that they are adapted precisely to that change of the living surrounding environment which called forth the injury. Such a division might appear artificial were we not dealing precisely with a historical regularity. But it is grounded in the fact that we can ascribe regularity only to those changes of form the conditions of which are laid down in the laws of motion of living matter itself. In this respect, consequently, adaptations based upon the utilisation of new sources of energy, and so forth, remain by their character and in the sense of the regularity of evolution always fortuitous, as conditioned by the states of non-living nature and their changes. On the contrary, those adaptations which represent direct adaptations to living nature and its changes bear the character of the historically necessary and are accompanied by an increase of external work.
Incidentally, this division corresponds to that which Severtsov draws in his book on the morphological regularities of evolution, when he speaks of aromorphoses and idioadaptations, finding what is historically regular in the direction, in the aromorphoses, leading to a constant intensification of all the vital functions. The relations between these two kinds of morphological changes and the regularity of the direction of evolution Severtsov formulates in the following manner: “The period of aromorphosis is characterised by the fact that during it there develop morphologico-physiological changes raising the intensity of the active functions, and idioadaptations not hindering this raising; the period of idioadaptation is characterised by the fact that during it there are formed only idioadaptations, altering the structure of the animals, but not raising the intensity of the vital energy: the level of vital energy and the complexity of structure remain, in these cases, in the descendants at the same level as in the ancestors”.[Author’s note:] A. N. Severtsov, Morphologische Gesetzmässigkeiten der Evolution, S. 156, Jena, 1931.
Returning to our theory of mutations and to the general theory of living matter, we see that what is historically regular in evolution will consist precisely in the alteration, namely in the increase, of µ₀, that is, of the potential of the egg cell, and not in the alteration of the fine structure of the chromosomes. For upon the latter depend the special hereditary structures conditioning idioadaptation, whereas upon µ₀ depends Rubner’s constant, that is, the whole energy transformed by a unit of living mass in the course of the life of the organism. But since the structures serving for external work likewise represent non-equilibrium structures of living matter, which are maintained at the expense of metabolism and must be renewed after each piece of work, it is clear that an increase of external work requires an intensification of metabolism, and consequently a raising of Rubner’s constant too. This connection becomes invalid only when the structures serving for external work are not living structures and therefore do not require internal work on the part of the organism for their maintenance, that is, when the organism employs machines, even if only in the most primitive form, such as for example levers — but with this there already begins humanisation, and there come into force the laws of motion of society, which thrust biological regularities into the background.
Thus, we arrive at the conclusion that in the course of evolution there takes place an increase of Rubner’s constant, and that this increase is conditioned not in a purely externally causal way, but is the result of a historical regularity, in its turn conditioned by the laws of motion of the whole of living matter on the Earth.
The fact of the increase of Rubner’s constant is illustrated by the table.
| t — duration of life in days | g — weight in kg | i — consumption of oxygen over 24 hours in calories | ||
|---|---|---|---|---|
| Coelenterata | ||||
| Carmanina hastata | 80 | 0.014 | 0.007 | 4×10 ⎫ |
| Rhizostoma pulmo | 80 | 0.077 | 0.083 | 7.7×10 ⎬ 5.8×10 |
| Echinodermata | ||||
| Cucumaria | 5×365 | 0.01 | 0.022 | 4×10³ 4×10³ |
| Vermes | ||||
| Hirudo | 20×365 | 0.010 | 0.008 | 5.8×10³ ⎫ |
| Lumbricus | 10×365 | 0.003 | 0.023 | 2.7×10⁴ ⎬ 1.5×10⁴ |
| Mollusca | ||||
| Limnaeus | 4×365 | 0.0023 | 0.022 | 1.4×10⁴ ⎫ |
| Ostrea virginica | 10×365 | 0.05 | 0.086 | 6.3×10³ ⎬ 1×10⁴ |
| Arthropoda | ||||
| Astacus | 25×365 | 0.031 | 0.137 | 4×10⁴ ⎫ |
| Palinurus | 25×365 | 0.25 | 1.27 | 4.6×10⁴ ⎬ 5.9×10⁴ |
| Homarus | 25×365 | 0.31 | 2.47 | 7.2×10⁴ ⎭ |
| Vertebrata | ||||
| Horse | 40×365 | 450 | 13260 | 4.3×10⁵ ⎫ |
| Sheep | 12×365 | 70 | 2856 | 1.8×10⁵ ⎬ 3.2×10⁵ |
| Dog | 11×365 | 15 | 1462 | 3.9×10⁵ |
| Cat | 9.5×365 | 2.5 | 204 | 2.8×10⁵ ⎭ |
| Human | 70×365 | 70 | 2550 | 9.3×10⁵ 9.3×10⁵ |
In this table there are, of course, still many gaps, and it is very inexact, but it nevertheless seems to us that it correctly conveys the actual tendency. It would be highly desirable to carry out systematic investigations of this magnitude and of its changes in the various species of the various groups of organisms.
It should be especially pointed out that the increase of Rubner’s constant by no means signifies a widening of the boundaries of physiological adaptation. An organism may possess a very small Rubner’s constant, while, with very strong fluctuations of some magnitude of the state of the surrounding environment, the actual number of calories formed by one gram of living substance may, within these boundaries, correspond to the magnitude of Rubner’s constant. On the contrary, with a large magnitude of Rubner’s constant, the quantity of calories actually formed may fall below this magnitude already with insignificant fluctuations of the surrounding environment. In other words, the material for evolution is supplied by organisms with narrow boundaries of adaptation, and not by those with a small Rubner’s constant. The result of the struggle for existence, however, is the survival of organisms with wider boundaries of adaptation or with a larger Rubner’s constant. But since organisms with wide boundaries of adaptation give less material, that is, fewer variants, the forms with wider boundaries of adaptation lag behind in evolution, and it is the forms with a larger Rubner’s constant that evolve. Thus what is historically regular will be not an increase of adaptability, but an increase of Rubner’s constant, whereas the adaptability to fluctuations of definite magnitudes of the state of the surrounding environment will reveal no regularity in its changes.
If we thus arrive at the conclusion that what is regular in evolution is the increase of external work and, in consequence of this, the increase of Rubner’s constant; if, further, we have drawn from our theory of living matter and of metabolism the conclusion that the increase of Rubner’s constant is conditioned by a raising of the potential of living matter — then, from the point of view of the fine structure of living matter, for which it is, after all, characteristic that it is built up out of deformed, active particles not corresponding to the equilibrium structure and subordinated to laws of motion maintaining precisely this state — from the point of view of this structure, we say, we may assert in a general form that evolution consists in the heightening and intensification of the fundamental properties of living matter.